- Details
- Zugriffe: 65467
Kapitel 06.04: Die Zellmembran und zelluläre Vorgänge
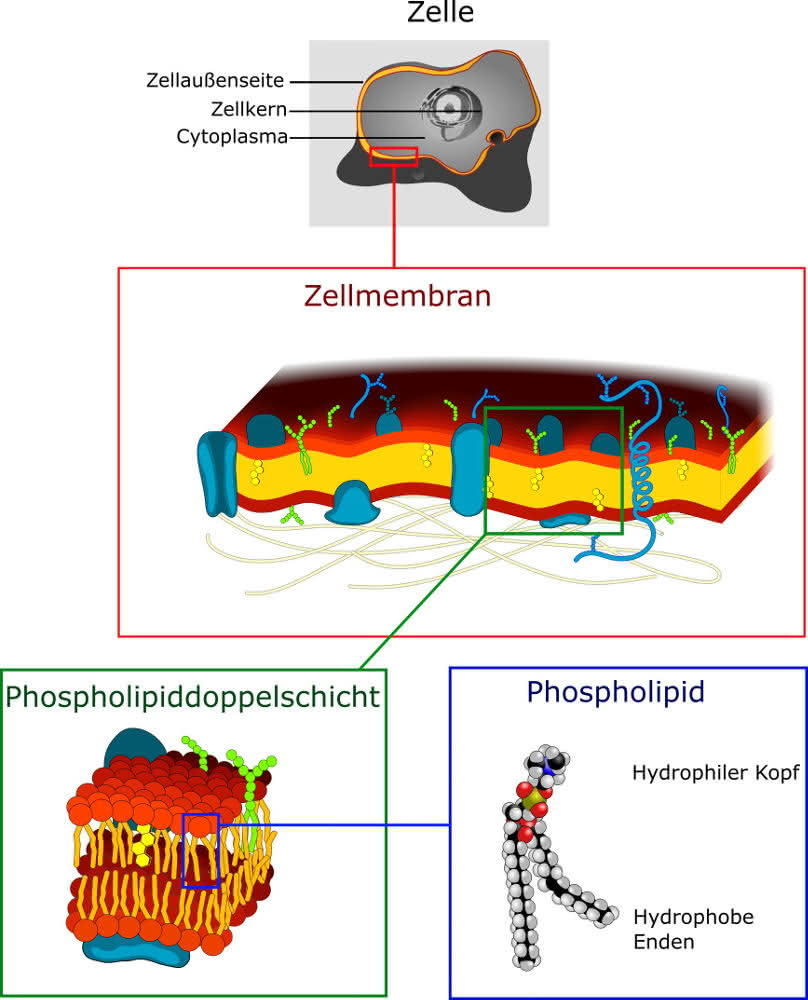
Quelle Bild: CC-By-SA 3.0 by Wikikommonsusers Dhatfield und LadyofHats, Marina Ruiz, Creative Commons Attribution-Share Alike 3.0 Unported; Thank You! https://commons.wikimedia.org/wiki/File:Cell_membrane_detailed_diagram_4.svg
Die Zellmembran
Jede Zelle, ganz egal ob tierische oder pflanzliche, ist umhüllt von einer Zellmembran. In diese Membran eingelagert ist eine Vielzahl verschiedenster Proteine, welche ihrerseits verschiedenste Funktionen erfüllen.
Viele sind Kanalproteine zum Durchtritt von z.B. Ionen oder anderer löslicher Substanzen. Andere wiederum stellen transmembranäre Rezeptoren für verschiedene Stoffe aus der Umgebung dar.

Dieses Bild einer Hefezelle wurde mittels Gefrierbruchverfahren präpariert und zeigt, wie dicht diese Proteine in der Membran gelagert sind. (Vergrößerung 22 000x)
Quelle Bild: EM-Aufnahmen mit freundlicher Genehmigung von Prof. Dr.
Biomembranen: Die Zellmembran
Zellen, aber auch deren Organellen sind von Biomembranen umgeben. Zellmembranen umschließen dabei das Cytoplasma und grenzen es nach außen ab. Zellmembranen sind sehr dünn und im Vergleich zur Zellwand viel weniger stabil.
- Die Zellmembran begrenzt das Plasma nach Außen (Plasmalemma).
- Bei Pflanzenzellen geht die Zellmembran über Plasmabrücken (Plasmodesmen) zwischen den Zellen kontinuierlich in die Membran der Nachbarzelle über. Solche Verbindungen dienen dem Stoffaustausch und der Kommunikation.
- Auch die viele Organellen, wie Vakuole und die Plastiden (Chloroplasten und Mitochondrien) sind von einer Membran umgeben (=Tonoplast)
- Membranen lassen Stoffe (z.B. Wasser und Salze) hindurch. Kleinere Moleküle oft passiv, größere aktiv (=> selektiv permeabel)
- Im Gegensatz dazu sind einfache Membranen nur selektivpermeabel, das heißt, sie sind für Wasser, das als Lösungsmittel fungiert, permeabel, für gelöste größere Teilchen (größere Moleküle und geladene Teilchen =Ionen) allerdings nicht.
- Die Außenseite der Zellmembran ist mit verschiedenen Rezeptoren und Oberflächenproteinen besetzt.
- Im Lichtmikroskop ist die Zellmembran meist nicht erkennbar, da die zwar elastisch verformbare und stabile Membran nur 6 bis 9nm dick ist.
Aufbau von Biomembranen:
In allen Biomembranen findet man als Bauelemente Lipide und Proteine und häufig geringe Mengen von Kohlenhydraten.
Mebranbestandteile: Lipide
- Lipide sind fette und fettähnliche Stoffe
- in der Biomembran findet man vor allem Phospholipide (Lipide, welche eine Phosphatgruppe enthalten) sowie Glykolipide (Lipide, welche Kohlenhydrate gebunden haben).
- Lecithin ist eines der häufigsten Membranproteine. Es ist ein Phospholipid.
- Lipide bilden eine Doppellipidschicht, deren polare (hydrophile) Enden dem wässrigen Umgebungsmilieu entgegenragen, während die apolaren, lipophilen Schwänze einander zugekehrt sind,
- Alle Lipide sind gut in Fett und organischen Lösungsmitteln löslich, aber unlöslich in Wasser
Lecithin - ein häufiges Membranlipid
Lecithin besteht als komplexes Molekül im Wesentlichen aus fünf chemischen Bestandteilen:
Cholin, Phosphat, Glycerin und 2 Fettsäuren.
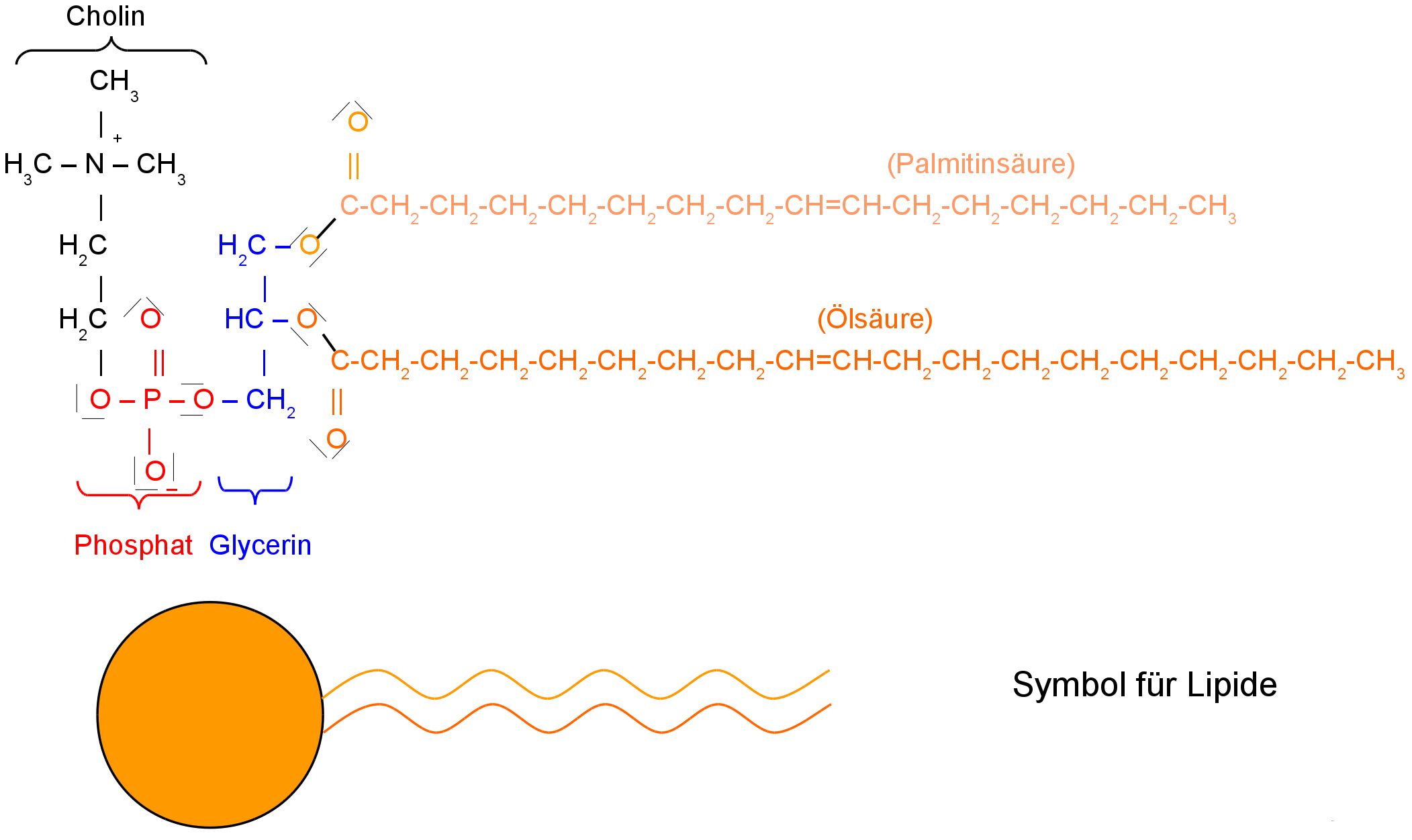
Phopshoglyzeride haben an das Glycrin sowohl Phosphat (PO4)3- als auch zwei Fettsäuren gebunden. Das Phosphoglycerid „Lecithin“ hat zusätzlich ein Cholin an das Phosphat gebunden.
Membranen bestehen aus einer Lipiddoppelschicht und enthalten Proteine
Man unterscheidet zwei Arten von Proteinen (=Eiweißen):
a) Periphere Proteine (=Oberflächenproteine) haften elektrostatisch von außen an der Membran
b) Integrale Proteinen sind in die Lipiddoppelschicht eingebunden und zum Teil auch von außen sichtbar. Sie können als Transportproteine (auch Tunnelproteine genannt) einen feinen Kanal bilden, der Ein- bzw. Austritt kleiner Moleküle und von Wasser ermöglicht.
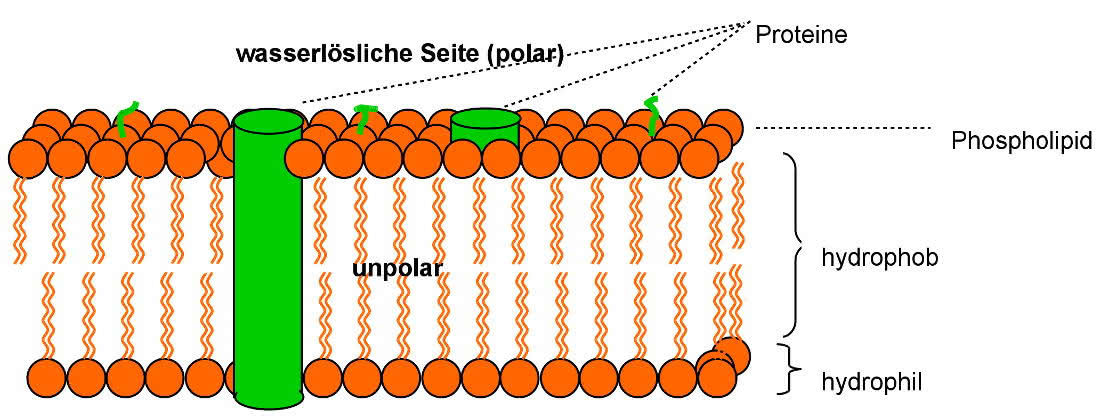
Die Umwandlungs- und Bewegungsvorgänge der Membran werden als Membranfluss bezeichnet, da alle Bestandteile zum selben System gehören und frei beweglich sind. Membranen sind nicht statisch und haben keine feste Struktur, sie können sich gegeneinander verschieben
Das Verhältnis von Lipiden zu Proteinen beträgt von 1:4 bis 4:1. Es ist abhängig von der Art der Stoffwechselaktivität der zugehörigen Zellen.
Die Zellmembran im Detail:
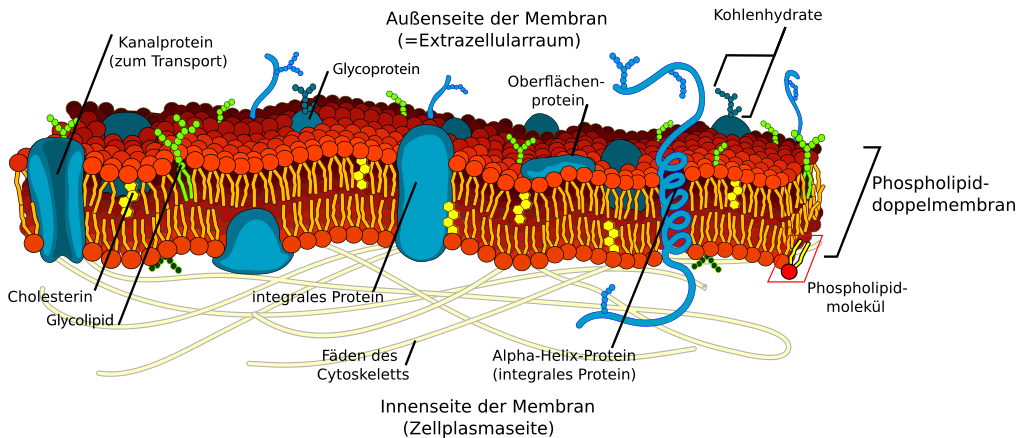
Quelle Grafik: Public domain nach der Grafik von Lady Of Hats, Marina Ruiz; https://commons.wikimedia.org/wiki/File:Cell_membrane_detailed_diagram_en.svg - Muchas Gracias
Aufgaben:
1. Diskutiere, inwiefern ein am Boden mit Murmeln gefülltes Gefäß Ähnlichkeit mit einer Membran hat?
2. Versuche ein besseres Membranmodell zu entwickeln.
Aufgaben von Membranproteinen
Membranproteine haben verschiedene Aufgaben. Sie werden von der Zelle speziell für ihren jeweiligen Einsatzzweck hergestellt:
- Transport von Stoffen durch die Membran: Tunnelproteine, Carrier, Kanalproteine
- Andockstellen für Stoffe: Rezeptorproteine (erkennen zum, Beispiel nach dem Schlüssel-Schlossprinzip Hormone, oder auch Fremdstoffe)
- Glycoproteine sind Membranproteine, welche Kohlenhydrate gebunden haben. Sie dienen zur Kommunikation zwischen Zellen (vor allem bei Immunzellen und Hormonrezeptorzellen) und machen einige Zelloberflächen gleitfähiger. In Membranen gibt es mehr Glycoproteine als Proteine, welche keine Kohlenhydrate gebunden haben.
Die Zellmembran ist eine Doppelmembran
Die Membran enthält viele Proteine. Diese durchziehen sie z.B. als Kanäle oder sind auf der Oberfläche zu finden. Zusätzlich befinden sich Glycolipide (phosphatfreie Membranlipide) und Cholesterinmoleküle an den Membranen. Zudem wird die Zellmembran von innen her durch Filamente (Fäden) des Cytoskeletts gestützt.
Die Membran ist als Doppellipidschicht aufgebaut. Sie ist dadurch außen hydrophil und innen hydrophob. Genauere Erklärungen im Kapitel zur Zellmembran.
Membranstrukturen zur Verbindung von Zellen
Desmosomen
Desmosomen sind sogenannte „Haftpunkte“, welche sich an Zellmembranen befinden. Sie halten vor allem die Zellen zusammen, welche sich in mechanisch stark beanspruchten Geweben (v.a. in Epithelien oder Muskelzellen) befinden. Auf das Gewebe wirkende Verformungsenergie, wie beim Dehnen und Zerren wird so gleichmäßiger verteilt und das Gewebe hält fester zusammen und reißt nicht. Die Haftplatte selbst hat eine stabilisierende Verbindung zum Cytoskelett.
Tight Junctions
Tight Junctions verbinden benachbarte Zellen mit Membranproteinfasern (Occludin &Claudin). Sie liegen wie Gürtel um Epithel- und Endothelzellen, so dass die Zellen fest zusammenhalten. Die wesentliche Auswirkung ist somit eine Einschränkung des Membranflusses, so dass andere Proteine an Ort und Stelle bleiben. Gleichzeitig wird so eine Diffusionsbarriere gebildet.
Gap Junctions
Gap Junctions sind tunnelartige Verbindungen zwischen Zellen, welche vor allem bei Nervenzellen, Sinneszellen und Muskelzellen auftreten. Durch sie können Moleküle schneller zwischen Zellen ausgetauscht werden, als durch die üblichen Membrantransportproteine. Zusätzlich können sie geschlossen werden.
Plasmodesmen
Bei Pflanzen ist das Zellplasma zweiter benachbarter Zellen nicht nur durch die Zellmembran voneinander getrennt, sondern zusätzlich durch die Zellwände jeder der beiden Zellen. Das macht einen Stoffaustausch schwierig und langsam. Plasmodesmen sind röhrenartige Verbindungen, die groß genug sind, dass selbst ER durch sie hindurch verlegt sein kann.
Diffusion durch Membranen
1. Neutralrotversuch im Reagenzglas:
In ein Reagenzglas wird Öl und Wasser gegeben und dann Neutralrot aufgelöst. Das überwiegend lipophile Neutralrot löst zu vor allem im Öl, welches eine rote Farbe annimmt. Das Wasser ist bestenfalls orange gefärbt. Nun gibt man Säure zu und das Wasser wird rot, wo hingegen sich das Öl entfärbt.
S: Der Farbstoff Neutralrot löst sich nicht gut in Wasser, da er ein fettlöslicher (=lipophiler) Farbstoff ist. Die Säure protoniert den lipophilen Stoff Neutralrot, wodurch dieser zum Neutralrotion wird (mit positiver Ladung durch das hinzugefügte Proton). Als Ion ist es hydrophil und löst sich in Wasser.
2. Neutralrotlösung wird zu Zwiebelzellen gegeben.
B: Die Vakuolen der Zwiebelzellen färben sich rot
S: Die Substanz Neutralrot kann als lipophile Substanz die Lipiddoppelschicht der Zellmembran und der Vakuolenmembran (Tonoplast) gut durchdringen. In der Vakuole herrscht aber ein leicht saure Milieu und es liegt Wasser vor. Nur wasserlösliche Stoffe können sich in der Vakuole lösen. Durch die Säure kommt es (wieder wie bei V1) zur Protonierung. Das Neutralrotion entsteht, welches sich gut im Wasser und dem hydrophilen Milieu der Vakuole auflösen kann. Die Vakuole färbt sich rot.
Der Farbstoff Neutralrot kann auch nicht mehr aus der Vakuole austreten, da Neutralrotionen hydrophil sind und nicht so leicht die lipophilen Schichten der Vakuolenmembranen durchdringen können.
Lipophile Stoffe können Membranen durchdringen. Hydrophile Stoffe können die Membran nur durchdringen, wenn sie sehr klein sind oder wenn es spezielle Ionenkanäle oder Carrier für sie gibt.
Beides gilt für den Stoff Neutralrot nicht. Es handelt sich um ein recht großes Molekül.
Die Vakuole ist für Neutralrot somit eine Ionenfalle geworden.
Aufgaben
- Das oben beschriebene Experiment zur Färbung der Vakuolen funktioniert nur, wenn man Leitungswasser verwendet. Ist entionisiertes (bzw. destilliertes) Wasser vorhanden, so färben sich nur die Zellwände (aber nicht die Vakuolen) rot. Das Eintreten des Farbstoffes findet nicht statt. Erkläre, warum das so ist.
Zur Erklärung dienen folgende Werte als Hilfe:
pH Leitungswasser > 8 -> basisch
pH destilliertes Wasser pH < 6 -> sauer)
Bestandteile der Zellwand wie Pektin tragen negative Ladungen.
Stofftransport durch Biomembranen
Betrachtet man Zellmembranen im Elektronenmikroskop, so sieht man, dass sich auf und in der Membran viele Proteine befinden. In einer Zelle können bis zu 100 000 verschiedene Proteinarten vorkommen. Sie ermöglichen vielfältige Reaktionen, wie z.B. den Transport von Stoffen in die Zelle.
Die Proteine sind allerdings nicht fest verankert, wie z.B. Pflanzen, die fest auf einer Mauer wachsen. Die Membran kann man sich eher wie eine Flüssigkeit vorstellen, in der die integralen Proteine „schwimmen“. Dabei können sie „Poren“ bilden.
Proteine befinden sich in und auf der Membran.
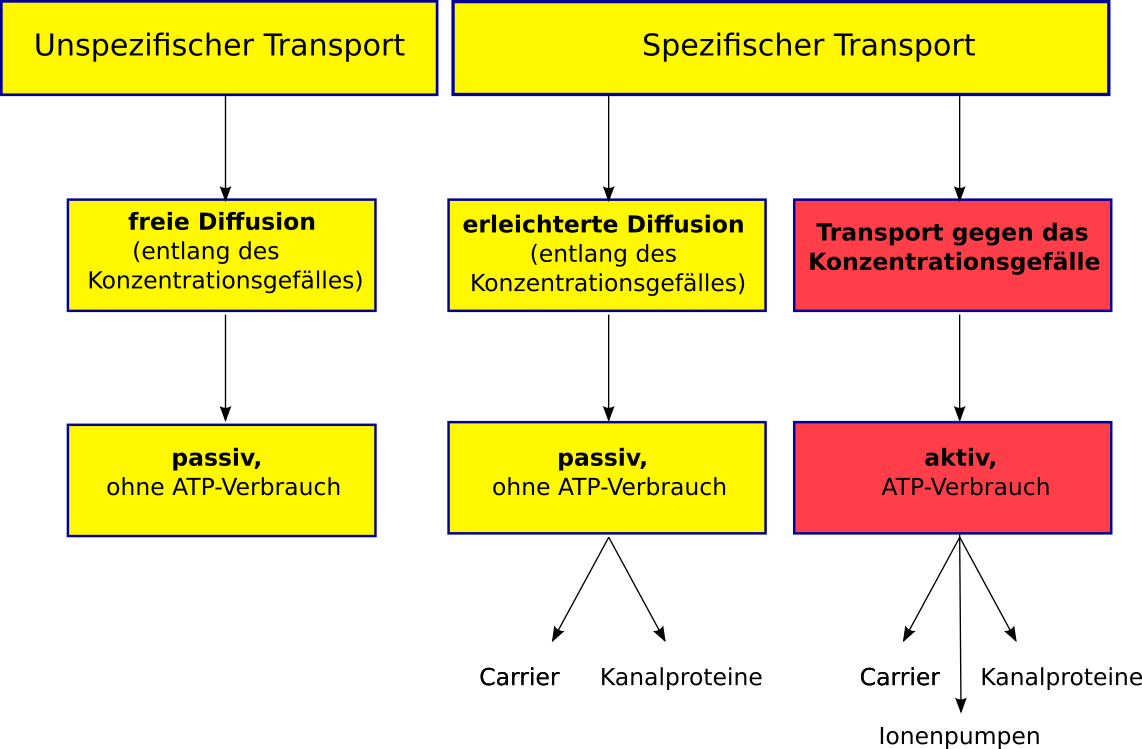
Stofftransport in und aus den Zellen: Es gibt nun mehrere Möglichkeiten für Stoffe, in die Zelle zu gelangen bzw. aus ihr heraus zu kommen. Einige laufen immer ab, andere werden von der Zelle genau nach Bedarf gesteuert.
1. Freie Diffusion:
Hinsichtlich der Durchlässigkeit kann man bei Membranen unterschiedliche Vorgänge beobachten:
Die freie Diffusion ist eine oft zu findende Möglichkeit des Eintritts in Zellen. Um sie zu verstehen, muss man wissen, dass in lebenden Systemen Membranen meist nur teilweise durchlässig (= selektiv permeabel) sind. Das bedeutet, dass die Biomembran für größere Moleküle (viele Salze, große Ionen, organische Stoffe wie Zucker usw...) ein Hindernis für den Eintritt in die Zelle darstellt. Diese großen Moleküle haben es wesentlich schwerer als kleine, da sie sich nicht durch die Phospholipide der Membran „durchmogeln“ können.
Kleine Teilchen, wie Wasser, Sauerstoff, Kohlenstoffdioxid und Glycerin können meist gut in die Zelle eintreten. Aufgrund der Eigenschwingung1 der Moleküle, ist es ihnen möglich, sich sozusagen durch die Lipiddoppelschicht hindurch zu zittern und so in die Zelle zu gelangen.
Wasser (H2O), Sauerstoff (O2), Kohlenstoffdioxid (CO2) und kleine Moleküle wie Glycerin (C3H5(OH)3) gelangen leicht in Zellen, indem sie die Biomembran einfach durchqueren.
Fettlösliche (=lipophile) Stoffe , wie z.B. das schädliche Lösungsmittel Benzol, gelingt dies auch, obwohl sie recht große Moleküle sind. Sie können sich sozusagen durch die Lipiddoppelschicht hindurchlösen. Dieser Vorgang ist vergleichsweise langsam.
Ein Vergleich zum Eintritt fettlöslicher Moleküle: Im Konzert stehen viele Menschen vor der Bühne. Kleinere Zuschauer können sich aber mit Beharrlichkeit durch die Menge bis zur ersten Reihen „drängeln“.
Aber was ist nun der Grund, warum Moleküle und Ionen überhaupt in die Zelle gelangen?
Die treibende Kraft ist alleine der Konzentrationsunterschied (also das Konzentrationsgefälle, auch Gradient genannt) zwischen dem Äußeren und dem Inneren der Zelle. Der Transport erfolgt freiwillig (also ohne Energiebedarf (ohne ATP-Verbrauch)) vom Ort hoher Konzentration zum Ort niedriger Konzentration. Dieser Ausgleich findet immer statt.
Der freiwillige Übertritt von Stoffen in Zellen hinein und aus Zellen heraus, wird freie Diffusion genannt. Die Zelle hat keine Möglichkeit, die Aufnahme und die Abgabe von Stoffen zu steuern. Energie wird dazu nicht benötigt!
Die Aufnahme und die Abgabe von Molekülen durch freie Diffusion hängt u.a. von der Größe und Ladung der aufzunehmenden Teilchen ab. Die Membran ist also selektiv permeabel.
Um z.B. die Wasserabgabe an heißen Sommertagen durch die Blätter zu verhindern, müssen die Zellen also eine andere Möglichkeit finden. Pflanzen haben zu diesem Zwecke oft eine wachsartige Isolationsschicht auf den Blättern, die Cuticula.
2. Spezifischer Transport durch Membranen:
Es gibt zwei Formen des spezifischen (gesteuerten und auswählenden) Transports:
a) Passiver Transport (=erleichterte Diffusion) durch Biomembranen:
Die Triebfeder bei der erleichterten Diffusion ist wie bei der freien Diffusion ein Konzentrationsunterschied zwischen der Innenseite und der Außenseite der Zelle. Der Unterschied liegt darin, wie Moleküle in die Zelle gelangen.
Durch bereits vorhandene Eintrittsöffnungen (dazu gehören Carrier (auch Translokatoren genannt) und Kanalproteine), gelangen selektierte Moleküle schneller in die Zelle als bei der freien Diffusion. Bei diesem Vorgang wird keine Energie verbraucht.
Wenn viele Moleküle eines Typs (z.B. die Ionen einer Salzlösung) vorliegen, geht diese Eintrittsmethode sehr schnell, da die Chance, das ein Ion den passenden Kanal trifft, recht hoch ist. Wenn aber sehr viele Moleküle vorliegen (also mehr Moleküle als Translokatoren vorliegen!) und alle Kanälchen besetzt sind, dann steigert sich die Eintrittsgeschwindigkeit nicht weiter, da alle Kanälchen „besetzt“ sind. Der „Sättigungswert“ ist erreicht.
Beispiel: der Ionentransport durch Nerven und Muskelzellen wird durch passiven Transport aufrechterhalten.
b) Aktiver Transport durch Biomembranen:
Nun ist es nicht immer so, dass Ionen vom Ort hoher Konzentration zum Ort der niedrigen Konzentration transportiert werden müssen. Gegen ein Konzentrationsgefälle (also vom Ort niedriger Konzentration zum Ort hoher Konzentration!) können (nur unter ATP-/Energieverbrauch) Moleküle auch transportiert werden. Dabei wird der Energieträger ATP in ADP + P zersetzt. Diese Zersetzung setzt Energie frei.
Auch hier werden dazu Carrier und Kanalproteine sowie die so genannten Ionenpumpen verwendet:
Spezifischer Transport durch Carrier
So genannte Translokatoren (=Carrier) sind tunnelartige Eiweiße, die sich in der Membran befinden und sie durchziehen. Man spricht von integralen Proteinen. Sie können Stoffe binden und durch die Membran hindurchbefördern. Dabei verändern sie ihre Form.
Vorteile dieser Methode:
- die Aufnahme ist schneller als bei der freien Diffusion
- die Carrier sind substratspezifisch, d.h. sie sind auf bestimmt Moleküle spezialisiert. Die Zelle kann so steuern, was sie aufnimmt.
Nachteil:
- beim aktiven Transport verbrauchen Carrier Energie in Form von ATP. Das ATP wird von den Mitochondrien gebildet und entsteht durch die Zellatmung, bei der z.B. Blutzucker mit Sauerstoff zu Wasser und Kohlenstoffdioxid oxidiert wird.
Transport durch Membranen
Schematische Anordnung möglicher integraler Membranproteine:
.png)
Quelle Grafik: Public domain nach der Grafik von Lady Of Hats, Marina Ruiz; https://commons.wikimedia.org/wiki/File:Scheme_facilitated_diffusion_in_cell_membrane-en.svg; https://commons.wikimedia.org/wiki/File:Scheme_secundary_active_transport-en.svg - Muchas Gracias
Man kennt drei verschiedene Carrier (spezifische Transporte)
Der Transport durch Membranen kann durch Translokatoren geschehen. Dies geschieht meist passiv, das heißt ohne dass Energie benötigt wird.
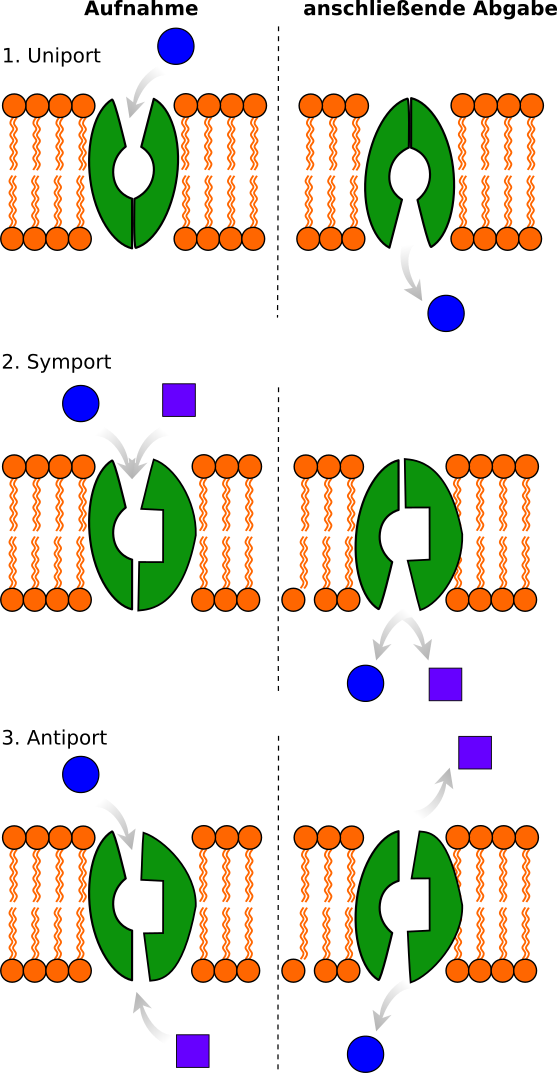
1. Uniport:
Ein Teilchen wird in nur eine Richtung durch
den Carrier/ das Kanalprotein (Translokator)
transportiert.
2. Symport
Zwei Substrate werden gleichzeitig, in eine Richtung
durch das Kanalprotein in eine Richtung transportiert.
3. Antiport
Zwei Substrate werden gleichzeitig in entgegengesetzte
Richtung transportiert. Eine Substanz gelangt in die
Zelle, die andere hinaus. Dazu ist meist Energie
in Form von ATP notwendig.
Endo- und Exocytose
Membranfluss:
Die Membran und ihrer Bestandteile sind kein starres, sondern ein dynamisches, bewegliches System. Es gibt viel Austausch zwischen den einzelnen Membransystemen. Die Umwandlungs- und Bewegungsvorgänge der Membran werden als Membranfluss bezeichnet, da alle Bestandteile zum selben System gehören und frei gegeneinander beweglich sind. Man kann sich vorstellen, dass die Phospholipide untereinander aber auch die Proteine aneinander vorbei gleiten können.
Endo- und Exocytose - Phänomene der Biologie:
Zellen geben Stoffe ab:
- Tränenflüssigkeit
- Verdauungsenzymen
- Speichel
- Schweiß
Zellen nehmen große Stoffe auf:
- Viren und Bakterien werden von Fresszellen aufgenommen
- Teer aus Zigaretten wird in der Lunge von Lungenzellen aufgenommen
- Einzeller nehmen Nahrungspartikel auf
Die Frage ist, wie Zellen so große Stoffe aufnehmen bzw. abgeben können, ohne dass sich die Zellmembran öffnet und die Zelle auslaufen würde. Eine Aufnahme über Kanalproteine/ Carrier ist ebenfalls ausgeschlossen, da diese Stoffe maximal in Molekülgröße aufnehmen können.
Exocytose (Abgabe von Sekreten aus Zellen):
Wenn nun Vesikel (=Membranbläschen) sich im Inneren der Zelle vom ER abschnüren und zu einem Golgi-Apparat werden, welcher dann wiederum Vesikel bilden kann (Golgi-Vesikel sind in der Regel mit Sekreten und Flüssigkeiten gefüllt), so können diese auch zur Zellmembran wandern und sich mit ihr vereinigen. Dies stellt kein Problem dar, da alle Membranen aus den gleichen Bausteinen bestehen und diese untereinander austauschbar sind.
Die Vesikel des Golgi-Apparates enthalten Sekrete (von Membranen umschlossen). Bei Kontakt mit der Zellmembran, wird die Vesikelmembran zur Zellmembran. Auf diese Weise kann dann die Flüssigkeit nach außen gelangen (=Exocytose).
Endocytose (Aufnahmevon Feststoffen und Flüssigkeiten in Zellen):
Beim umgekehrten Vorgang schnüren sich Vesikel von der Zellmembran ab, die sich ins Zellinnere bewegen. Werden dabei feste Partikel (z.B. Pflanzenreste, Bakterien, Stärkekörner) aufgenommen, spricht man von Phagocytose (phagein, gr.= fressen; cytos, gr.=Zelle). Bei Flüssigkeiten nennt man den Vorgang auch Pinocytose.
Ein dritter Vorgang ist die rezeptorgesteuerte Aufnahme. Dazu sitzen an der Membran bestimmte Rezeptoren, welche bei Kontakt mit bestimmten (gewünschten) Stoffen, die Einstülpung der Membran auslösen. Die entstehenden Vesikel haben eine raue Oberfläche, da die Proteine sich noch darauf befinden. Man nennt sie „Coated Vesikels“.
Durch die rezeptorgesteuerte Endocytose werden besonders Flüssigkeiten aufgenommen.
Alle drei Vorgänge der Endocytose finden an vielen Membranabschnitten statt (=> viele Einschnürungen). So können beispielsweise Amöben durch Phagocytose kleinere Einzeller als Nahrung aufnehmen. Beim Pantoffeltierchen ist ein spezielles Feld, das so genannte Mundfeld, für die Nahrungsaufnahme durch Phagocytose vorhanden. Im Darm wird auf diese Weise auch Fett resorbiert (=aufgenommen).
Übersicht - Endo- und Exocytose
Durch Vorgänge wie Endo- und Exocytose wird der Membran Material weggenommen oder wieder hinzugefügt. Die Membran ist somit immer im ständigen Wandel und Umbau!
Membranen sind somit nicht statisch anzusehen, sondern vielmehr immer in Bewegung und „fließend“.

Beispiele für Exocytose:
a) an den Enden der Axone der Nervenzellen befinden sich die Endknöpfchen. Sie geben z.B. durch Exocytose Neurotransmitter in den synaptischen Spalt.
b) Abgabe von Sekreten aus Drüsen, z.B. Speicheldrüse, Tränendrüse, Schweißdrüse; Bauchspeicheldrüse usw...
Lysosomen
Kleine Vesikel, in denen Stoffe umschlossen sind, werden als Vesikel bezeichnet. Es können dabei zum Beispiel Produkte des Golgi-Apparates umschlossen sein, oder auch von Einzellern durch Endocytose aufgenommene und dann durch Membran umschlossene Nahrung sein.
Warum schrumpeln die Hände beim Baden?
Beim Schwimmen, Baden und beim Abwaschen wird nach einiger Zeit die Haut schrumplig. Ursache ist einerseits eindringendes Wasser durch passive Membranvorgänge n die Zellen.
Unsere Zellen sind an den Fingern und Zehen aber andererseits auch stark von bereits abgestorbener Hornhaut bedeckt. Sie enthält sogenannte Hornzellen. Die Hornhaut nimmt auch Wasser auf! Sie saugt das Wasser zwar nicht ein, aber es kommt zum Aufquellen der Hornhaut durch das Eindringen in die Zellen und Zwischenzellräume. Die Hornhaut ist aber noch teilweise an die darunterliegende Unterhaut gebunden. Durch die nicht konstante Verbindung ist das Quellen aber nun nicht regelmäßig.
Da nur Hände und Füße bis zu 40 Lagen an Hornzellen enthalten, sieht man die Verschrumpelungen auch nur an diesen Stellen.
Vorversuche zum tieferen Verständnis der Diffusion
V: Eine Petrischale wird auf den Overheadprojektor gestellt und dann ein eines Körnchens Kaliumpermanganat in Wasser hnzugegeben.
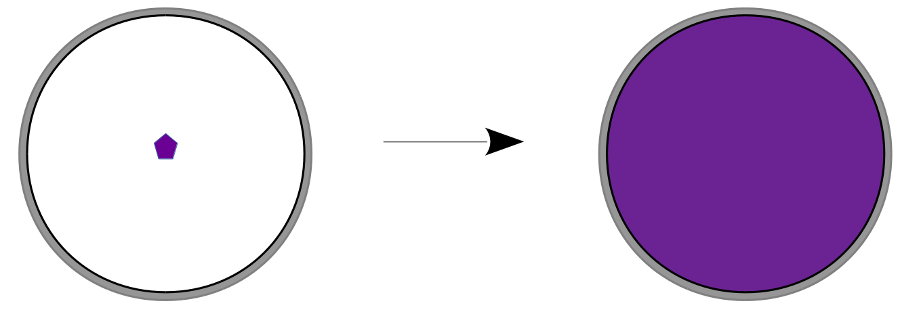
B: Der Farbstoff verteilt sich scheinbar von selbst. Nach einiger Zeit ist er im ganzen Gefäß fein verteilt.
S: Durch die Brown'sche Molekularbewegung (=eigene Zitterbewegung aller Teilchen!) stoßen Teilchen immer wieder aneinander und verteilen sich so im zur Verfügung stehenden Raum. Es kommt zu einem ungerichteten Konzentrationsausgleich.
Je wärmer dabei eine Lösung ist, desto stärker ist die Eigenbewegung der Teilchen.
Osmose & Diffusion
1. Volumen- und Gewichtsänderung eines Hühnereies in verschiedenen Lösungen
Unter der Kalkschale des Hühnereis befindet sich eine selektivpermeable Membran (=Membran, die nur in eine Richtung durchlässig ist). Um diese Membran zu untersuchen, wird die Kalkschale entfernt.
V: Die Kalkschale des Eies wird entfernt, indem es für ca. 6h in 10% Essigsäure gelegt wird. Das Ei wird dann abgewogen und in destilliertes Wasser (=hypotonische Lösung) gelegt.
Nach ca. 20 min. wird das Ei erneut abgewogen und anschließend in eine gesättigte Kochsalzlösung (hypertonische Lsg.) gelegt. Nach ca. 20 min. wird das Ei erneut gewogen.
B:
-
Lösung
Gewicht
hypotonisch (destilliertes Wasser)
schwer
„normal“ Zustand
normal
Hypertonisch (Salzlösung)
leichter
Schlussfolgerung: (siehe Erklärung bei dem folgenden Versuch mit den Zwiebelzellen.)
2. Versuch zu Plasmolyse - Deplasmolyse:
Als Material dienen Zellen einer roten Zwiebel (aus der äußeren einschichtigen Epidermis einer Zwiebelschale) oder Zellen aus dem Fruchtfleisch von Ligusterbeeren. Die Ligusterzellen erhält man durch Abschaben von etwas Fruchtfleisch mit dem Skalpell
|
Versuch |
Beobachtung |
|
1. Die Zellen des Zwiebelhäutchens werden auf einen Objektträger gebracht, ein Deckglas darüber gelegt und im Mikroskop beobachtet
|
Die rötlich gefärbten Zellsaftvakuolen nehmen fast das gesamte Zellvolumen ein. |
|
2. An den Rand des Deckglases wird eine hypertonische Lösung (z.B. konzentrierte Salzlösung) getropft und mit etwas Filterpapier unter das Deckglas gesaugt.
|
Die Zellsaftvakuole verkleinert sich und färbt sich dadurch dunkler. Das Schrumpfen der Vakuole erfolgt unmittelbar nach dem Durchsaugen der Kochsalzlösung. |
|
3. Nun werden einige Tropfen destillierten Wassers auf den Objektträger gebracht und unter das Deckgläschen gesaugt |
Die Zellsaftvakuolen haben nach kurzer Zeit wieder die ursprüngliche Größe und Färbung. |
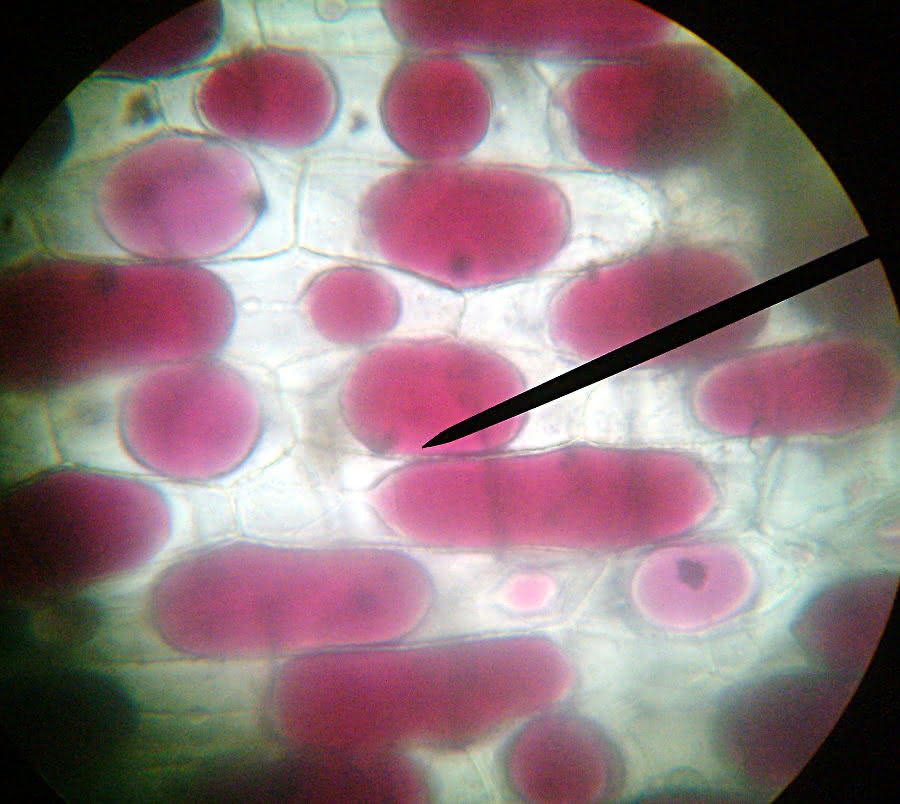
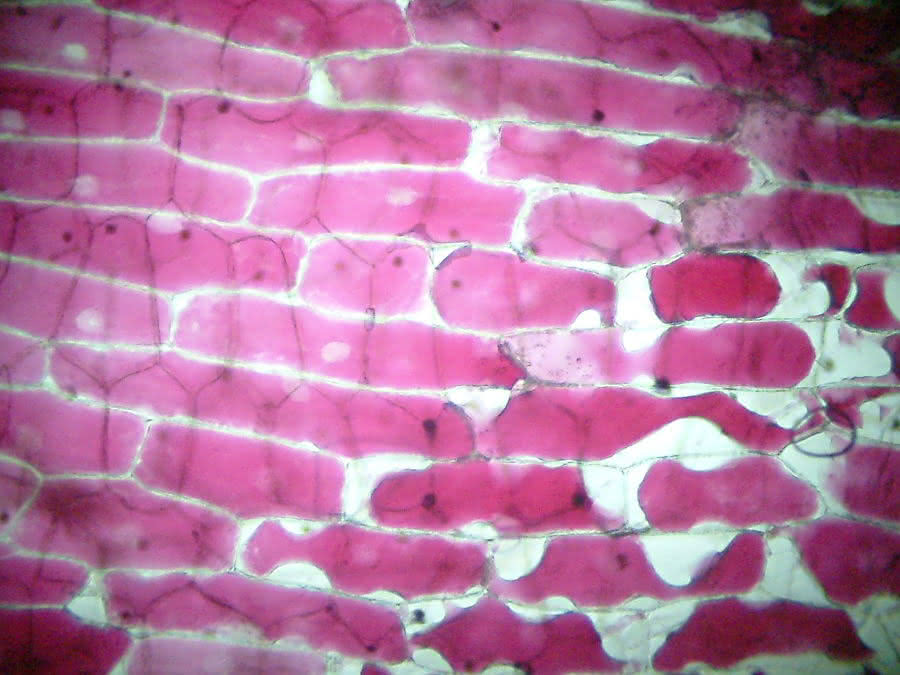
Zwiebelzellen vor Zugabe des Salzwassers:

Plasmolyse - isotonische Lösung
Zwiebelzellen kurz nach Zugabe d. Salzwassers:
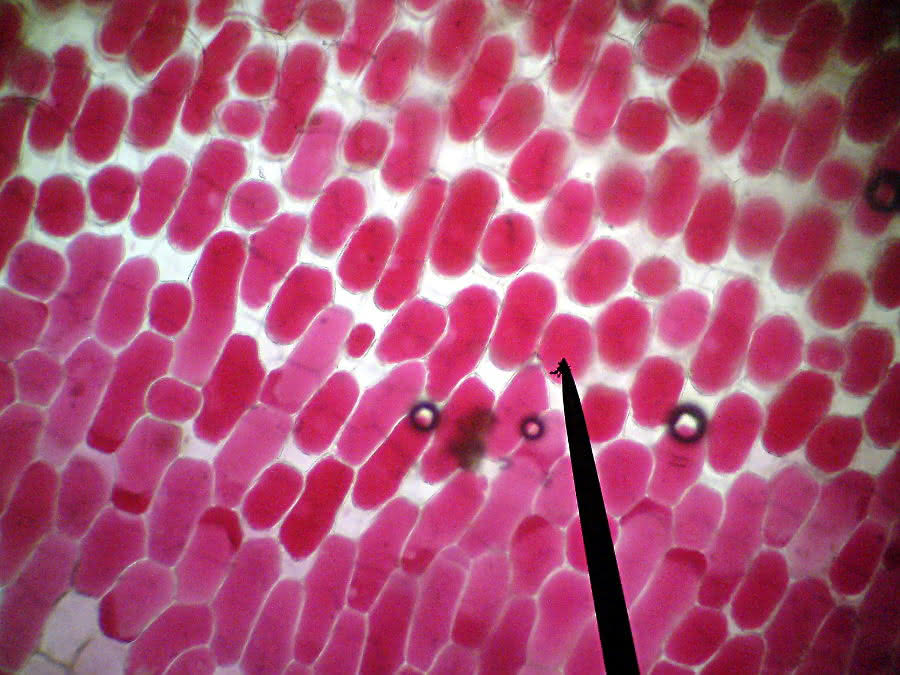
Vergrößerung:
Was schätzt Du, von welcher Seite das Salzwasser zugefügt wurde und warum nicht alle Zellen eine Plasmolyse zeigen?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Grenzplasmolyse
In diesem Zustand ist keine Umkehr mehr möglich! Die Zelle hat zu viel Wasser verloren, die Vakuolen reißen oft ein, sind beschädigt und verlieren ihre Funktion. Die Zelle stirbt ab!
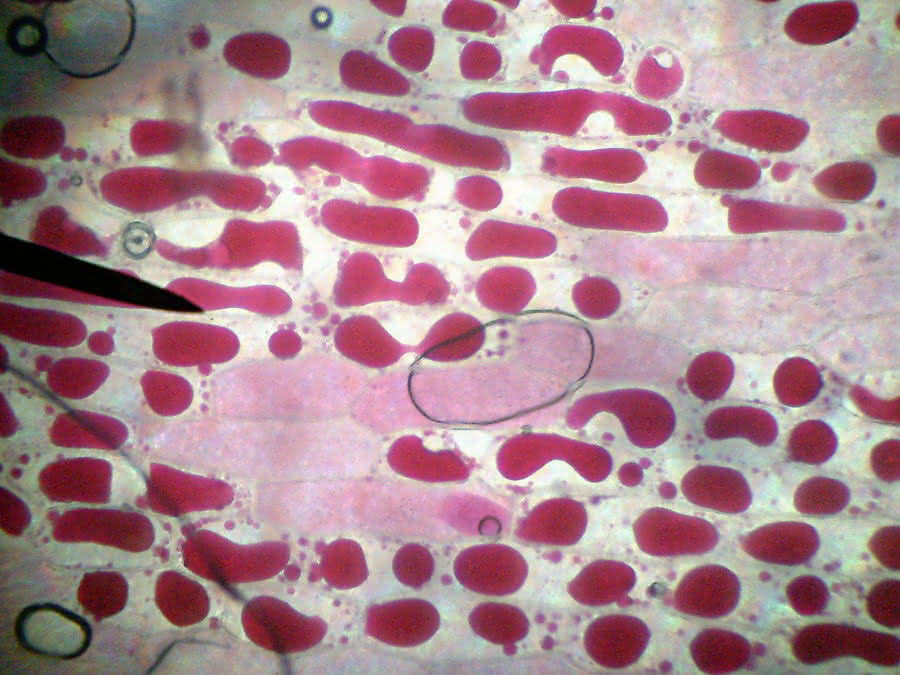
Grenzplasmolyse
Schlussfolgerungen: Um die Vorgänge zu verstehen, muss man wissen, was Diffusion ist:
Die Diffusion ist ein Vorgang, bei dem sich Atome oder Moleküle in Flüssigkeiten oder Gasen frei ausbreiten. Grundlage der Diffusion ist die Brown'sche Molekularbewegung (=Eigenbewegung von Atomen und Molekülen).
Aber, in Zwiebelzellen kann keine freie Diffusion stattfinden, denn die Membran behindert die freie Diffusion. Lebende Zellmembranen sind selektiv permeabel (ein etwas älter Ausdruck lautet auch semipermeabel) für Wasser und kleinere Ionen. Das bedeutet, dass sie für einige Bestandteile wie Wasser und kleine Ionen durchlässig ist, für große Ionen aber nicht!
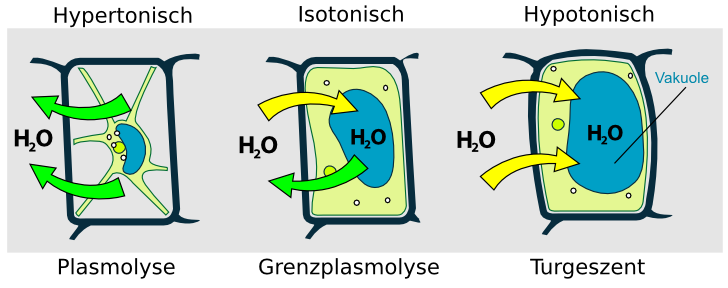
Bringt man eine lebende Zelle in eine Lösung, in der außen eine höhere Konzentration an Ionen als in der Zellsaftvakuole herrscht, so diffundiert Wasser aus der Vakuole durch die Zellmembran in die umgebende Lösung (da jetzt mehr Wassermoleküle von innen auf die Vakuolenmembran prallen als von außen).
Die mit dem Wasseraustritt verbundene Volumenabnahme der Vakuole führt zunächst zu einer Abnahme des Zellturgors (=Wanddruck die Zelle gegen die Zellwand - der Druck wird ausgelöst durch eine prall mit Flüssigkeit gefüllten Vakuole), dann zu einem Ablösen des Protoplasten von der Zellwand (=Plasmolyse). Dieser Vorgang ist reversibel (die Umkehrung heißt Deplasmolyse).
Quelle Bild: Public domain by wikicommonsuser LadyofHats, Marina Ruiz - Muchas Gracias; https://commons.wikimedia.org/wiki/Media:Plasmolyse_Pflanzenzelle.svg
{kind=link}
Unter Plasmolyse versteht man das Ablösen der Vakuolenmembran von der Zellmembran. Ursache ist die mit Wasseraustritt verbundene Volumenabnahme der Vakuole. Sie entsteht durch eine höhere Außenkonzentration an Ionen im Vergleich zur Konzentration in der Zelle.
Die Deplasmolyse ist der umgekehrte Vorgang!
So diffundiert Wasser aus der Vakuole durch die Zellmembran in die umgebende Lösung.
Die „Antriebsfeder“ der Plasmolyse ist der Konzentrationsunterschied zwischen der Innen- und der Außenseite. Die Plasmolyse ist auf lebende Zellmembranen beschränkt.
Unter Osmose versteht man den Vorgang der Diffusion eines Lösungsmittels durch eine selektivpermeable (=teildurchlässige) Membran.
Achtung: In einigen Büchern ist die Plasmolyse als die Wasserbewegung beschrieben. Dies ist aber im Grunde nur die Osmose bzw. das osmotisches Eindringen/ Austreten des Wassers.
Voraussetzung für die Osmose ist, dass auf beiden Seiten der Membran zwei Lösungen unterschiedlicher Konzentration (bzw. eine Lösung und das reine Lösungsmittel, z.B. Wasser) angrenzen. Von der Seite der stärker konzentrierten Lösung prallen im Durchschnitt weniger Lösungsmittelmoleküle auf die selektivpermeable Membran als von der weniger stark konzentrierten Lösung. Als Folge durchdringen viel mehr Moleküle der stark konzentrierten Lösung die Membran, als umgekehrt, von der schwach konzentrierten Lösung her in die stark konzentrierte übergehen.
3. Versuch zur selektiven Permeabilität lebender Membranen:
V: 3 Kartoffelhälften werden innen ausgehöhlt, so dass eine ca. 1cm tiefe und 1cm breite Vertiefung entsteht. Eine der Kartoffelhälften wird einige Minuten gekocht. In die Vertiefung der gekochten Kartoffel und in eine Vertiefung der rohen Kartoffel wird Kochsalz gefüllt. Die zweite rohe Kartoffel bleibt ohne Salzzugabe. Die drei Kartoffelhälften werden in eine mit Wasser gefüllte Petrischale gebracht.
Beobachtung:
Nach ca. 30-40 Min. ist die Vertiefung der rohen Kartoffel, die Salz enthielt, mit Wasser gefüllt. Bei der gekochten Kartoffel hat sich kein Wasser in der Vertiefung angesammelt, ebenso wie bei der rohen Kartoffel ohne Salzzugabe.
Schlussfolgerung:
1. In der rohen Kartoffel wird durch das Salz aus den umliegenden Zellen osmotisch Wasser entzogen.
2. In der rohen Kartoffel ohne Salzzugabe hingegen findet kein Wasseraustritt aus den Zellen der Kartoffel statt.
3. Die Beobachtung, dass sich in der gekochten Kartoffel trotz des Vorhandenseins von stark konzentriertem Salz kein Wasser in der Vertiefung ansammelt, zeigt die Beschränkung der selektiven Permeabilität lebender Zellen. Durch das Kochen werden die Proteine der Zellmembran zerstört und damit auch die Kanäle zum Wasseraustritt aus der Zelle.
4. Versuch: „Kartoffel - Osmometer“
Das „Kartoffel-Osmometer“ ist ein einfaches, anschauliches Beispiel eines Osmometers, wie es nach Stocke aufgebaut ist (Stocke`sches Gerät mit Pfeffer'schen - Zelle).
Zum Aufbau wird in eine Kartoffel mit einem Korkbohrer ein Loch gebohrt, dieses mit Salz gefüllt und mit einem durchbohrten Gummistopfen verschlossen. In das Loch des Gummistopfens wird ein dünnes Steigrohr gebracht. Diese „Apparatur“ wird nun in Wasser getaucht.
Erklärung: Die hohe Salzkonzentration im Inneren der Kartoffel ist stark osmotisch wirksam und entzieht dem Kartoffelgewebe Wasser. Das Gewebe wiederum nimmt aus der Umgebung Wasser auf. Die Aufnahme von Wasser wird im Steigrohr sichtbar. Das Wasser steigt solange, bis der hydrostatische Druck der Wassersäule im Steigrohr den Effekt des Konzentrationsgefälles kompensiert. Das Kartoffelgewebe entspricht der selektivpermeablen Membran einer Pfeffer‘schen - Zelle.
Zusammenfassung Osmose:
Grundsätzlich findet in der Natur immer ein Konzentrationsausgleich vom Ort der hohen Konzentration zum Ort der geringen Konzentration statt. Liegt eine selektivpermeable Membran vor, welche nur für Wasser durchlässig ist, so kann zwischen den gelösten Stoffen auf beiden Seiten der Membran kein direkter Konzentrationsausgleich stattfinden!
Stattdessen durchdringen Wassermoleküle die Membran und sorgen so für eine Verdünnung auf Seiten der hohen Konzentration. Auch dies führt zum Ausgleichen des Konzentrationsunterschiedes.
Diese Bewegung der Wasserteilchen findet man in vielen Bereichen der Natur:
Essen Menschen salzige Nahrung, gleicht der Körper dies mit Körperwasser aus, und dehydriert dabei. Er bekommt Durst und muss neues Wasser aufnehmen.
Aufgaben:
1. Erkläre die folgenden Begriffe: Plasmolyse, Turgor, selektivpermeabel, Osmose, Diffusion
2. Seeleute wurden früher davor gewarnt, in Seenot, selbst wenn sie am Verdursten waren, Meerwasser zu trinken. Kannst erklären, warum?
3. Sportler warnen: Nach dem Sport sollte man keinen starken Kaffee trinken (zumindest nicht ausschließlich!), da dieser den Körper dehydriert. Stelle die Verbindung von Wasserbedarf durch Leistungssport und Kaffeegenuss dar.
Die Osmose als Triebfeder der Plasmolyse
Am Anfang befindet sich außerhalb von Zellen oder Gewebe eine hoch konzentrierte Salzlösung. Als Folge löst sich bereits nach kurzer Zeit der Protoplast (=lebender Teil der Zelle) von der Zellwand ab. Die Vakuole wird kleiner, bis sich der Protoplast vollständig von der Zellwand löst und abkugelt.
Dabei hängt die Stärke des Zusammenziehens des Protoplasten maßgeblich von der Konzentration der verwendeten Salzlösung ab!

Ursache:
Das Wasser innerhalb der Zelle ist weniger stark konzentriert, als das Wasser außerhalb. Derartige Ungleichgewichte gleichen sich in der Natur (wenn möglich!) automatisch aus. Wasser diffundiert also aus dem Zellplasma und der Zellsaftvakuole hinaus. Die Triebfeder dieses hier stattfindenden Konzentrationsausgleiches sind die unterschiedlichen Konzentrationen an gelösten Salzen zwischen der Lösung außen und innen.
Der Ausgleich kommt nun vor allem durch das Diffundieren von Wasser durch die Zellmembran zustande. Dies ist alles nur möglich, weil Biomembranen vor allem für Wasser, aber nicht für gelöste Stoffe durchlässig sind. Man sagt, die Membran ist selektiv permeabel.
In der Folge wird also die hypotonischere (also weniger gelöste Teilchen enthaltene) Lösung im Inneren der Zelle mit der hypertonischen (also mehr gelöste Teilchen enthaltene) Lösung außerhalb der Zelle ausgeglichen.
Die stattfindende Diffusion von Wasser durch eine selektivpermeable Membrannennt man auch Osmose.
Ein Vergleich zum Verständnis:
Wenn man Wasser im Kochtopf verdampft, bildet sich ein Kalk(salz)belag am Boden.
Wenn man kurz vorher aber den Topf schon vom Herd nimmt, ist das restliche Wasser sehr viel salzhaltiger als das Ursprungswasser.
=> Die Wasserabgabe der Zelle (bei der Plasmolyse) erhöht in Wirklichkeit die Salzkonzentration innerhalb der Zelle, welche sich somit dem Außenmedium in der Konzentration angleicht (und dieses wird ja zusätzlich durch das aus der Zelle strömende Wasser verdünnt)
Zusatzinformationen:
https://de.wikipedia.org/wiki/Plasmolyse
Turgor und Wanddruck
Eine Zelle im Normalzustand hat einen Wanddruck von = 0. Das bedeutet, dass die Saugspannung gleich dem osmotischen Wert des Zellplasmas ist.
Bei Wasseraufnahme ist der Wanddruck größer als 0, die Saugspannung wird folglich sinken (da die Verdünnung im Inneren immer weiter zunimmt)!
Das geht soweit, bis die Saugspannung durch weitere Wasseraufnahme immer geringer wird, solange bis die Zelle kein Wasser mehr aufnehmen kann. Diesen Zustand nennt man „turgeszent“.
Die osmotische Zustandsgleichung für Zellen lautet: S = O - W
S = Saugkraft;
O = osmotische Wert;
W = Wanddruck ist
Aufgaben zur Osmose und zur Plasmolyse
1. Blutkörperchen müssen, wenn sie z.B. unter dem Mikroskop beobachtet werden in einer sogenannten isotonischen Kochsalzlösung sein. Kannst Du mithilfe der Grafik erklären warum?
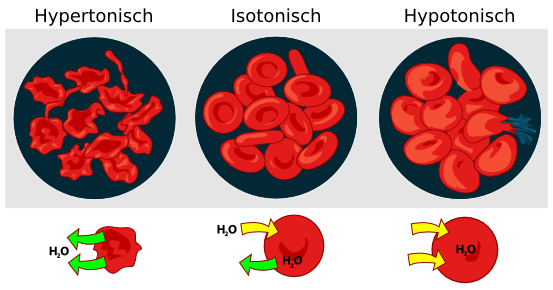
Quelle Bild: Public domain von Wikicommonsuser LadyofHats, Marina Ruiz Muchas Gracias; https://commons.wikimedia.org, /wiki/File:Osmotic_pressure_on_blood_cells_diagram.svg
{kind=link}
2. In Süßwasser lebende Pantoffeltierchen haben pulsierende Bläschen, Wimperntiere, welche im Meer leben, haben diese nicht. Welche Ursachen hat das?
3. Pflanzen am Straßenrand sterben oft ab, wenn im Winter Streusalz verwendet wurde. Nenne und erkläre die Ursachen.
4. Eine Wiese wird versehentlich zweimal in kurzer Zeit mit Mineralsalzdünger gedüngt. Der Rasen bekommt helle Flecken und scheint zu vertrocknen. Erkläre!
5. Regen bringt reife Kirschen zum Platzen. Begründe, warum das so ist.
6. Starker koffeeinhaltiger Tee (und auch Kaffee) mit Zucker führen zu mehr Harndrang, als wenn man die gleiche Menge Wasser getrunken hätte. Erkläre!
Aufnahme von Giftstoffen in das Gewebe
Aufnahme von Giftstoffen in den menschlichen Körper erfolgt durch die Zellen der Haut, besonders durch Inhalation von Dämpfen, Kontakt mit der Haut, Verschlucken oder bewusste Aufnahme. Die Giftstoffe gelangen je nach Weg zuerst meist in die Lunge, das Blut, die Nieren und den Darm.
Es folgt eine kleine Auswahl von Schadstoffen. Einige sind mittlerweile in ihrer Verwendung eingeschränkt oder verboten. Andere fügt man sich zum Teil bewusst zu.
- Pflanzenschutzmittel
- Kohlenwasserstoffverbindung (vor allem die chlorierten Kohlenwasserstoffe)
- DDT
- Schwermetallverbindungen
- Formaldehydausdünstungen
- Schimmelpilzgifte
- Verbrennungsrückstände (verbrannte Bratwurst!)
Die Aufnahme in die Zellen erfolgt über die bisher gelernten Mechanismen.
Alle folgenden Stoffe haben gemeinsam, dass sie die normale Funktionvon Zellen beeinflussen und somit schädlich sind.
Weitere Informationen zu diesem Thema kannst Du im Kapitel „Drogen“ nachlesen!
Untersuchungsmethoden von Zellmembranen
a) Elektrophorese:
Auf einer Gelplatte wird eine Untersuchungssubstanz (z.B. ein Lipid) aufgetragen und an beiden Enden der Gelplatte wird eine Gleichspannung angelegt. Die polaren Bestandteile werden zu den entsprechenden Polen gezogen, die apolaren bleiben in der Mitte. (Die Elektrophorese wird auch zur Untersuchung von Proteinen benutzt.)
b) Fettnachweise
- Fettflecknachweis
- Dünnschichtchromatographie von Lipiden
- chemischer Nachweis (mit Sudan-(III)-Glycerin)
Zu Fetten wird Sudanrot als Feststoff zugefügt der Farbstoff, wird von Fetten adsorbiert , sie färben sich rot. Fette extrahieren den Farbstoff aus seinen wässrigen Lösungen
c) Elektronenmikroskopische Nachweise - TEM (Transmissions Elektronen Mikroskop)
Die Auflösungsgrenze des Auges liegt bei etwa 0,1mm. Mit dem Lichtmikroskop lässt sich eine maximale Auflösung von etwa 500nm erreichen (1 nm = 1/1000mm), Mit einem TEM , welches anstatt eines Lichtstrahls einen Elektronenstrahl & anstatt optischen Linsen elektromagnetische Linsen besitzt) liegt die Auflösung ca. bei 0,3nm.
Betrachtet man die Vergrößerung so kann mit dem Lichtmikroskop eine etwa 2000fache und mit einem TEM ca. eine 1000000 Vergrößerung erreichen. Der Grund dafür liegt in der Wellenlänge, die beim sichtbaren Licht mindestens bei 380nm liegt, aber bei Elektronen (bei 100kV) nur 0,0038nm beträgt.
Allerdings kann man beim TEM im Vergleich zum Lichtmikroskop keine lebenden Objekte betrachten. Die Proben müssen zuerst also speziell präpariert werden, entweder durch chemische Fixation oder durch Einfrieren (Cryopräparation). Zudem müssen die Objekte bei der TEM sehr dünn sein (max.100nm).
d) Elektronenmikroskopische Nachweise - REM (Raster Elektronen Mikroskop)
Das REM funktioniert nach einem ähnlichen Prinzip, bei dem ein Elektronenstrahl über das Objekt geführt wird und Wechselwirkungen der Elektronen mit dem Objekt zur Erzeugung eines Bildes vom Objekts genutzt werden. Der maximale theoretische Vergrößerungsfaktor liegt etwa bei 1:500.000.
Das Rasterelektronenmikroskop basiert auf der Abrasterung der Objektoberfläche mittels eines fein gebündelten Elektronenstrahls. Der komplette Vorgang findet normalerweise im Hochvakuum statt, um Wechselwirkungen mit Atomen und Molekülen in der Luft zu vermeiden.
Der Elektronenstrahl wird in einer Feldemissionselektronenkanone erzeugt. Dabei kommen z.B. Drähte aus Wolfram zur Verwendung, welche erhitzt werde und dabei Elektronen abgeben. Dieser Elektronenstrahl wird mit Hilfe von Magnetspulen auf einen Punkt auf dem Objekt fokussiert.
e) Myelinfiguren
V & B: Öl und Wasser (die Wasserphase kann gefärbt werden) werden vermischt. Es bilden sich zwei Phasen. Dann fügt man Lecithin hinzu. Es bildet sich eine Emulsion
S: Es gibt Stoffe, die sich nicht miteinander mischen lassen. Fette sind lipophil und schwimmen auf der wässrigen Schicht. Diese wird lipophob oder auch hydrophil genannt. Erst die Zugabe eine Emulgators (wie hier dem Lecithin), können sich beide Phasen miteinander vermischen. Die vermittelnde Wirkung des Emulgators findet nur an den Grenzflächen statt. Aus diesem Grunde hilft ein gutes Schütteln, da so viele Grenzflächen entstehen. Dadurch färbt sich sie Emulsion meist weiß-trüb.
Beobachtung eines Ölflecks auf einer Wasseroberfläche
V: Fülle eine Petrischale mit Wasser und gib einen Ölfleck auf die Oberfläche. Was passiert, wenn Du etwas Spüli zufügst?
Was ist der Unterschied zwischen Fettsäuren, Fetten und Lipiden?
Lipid ist eine Sammelbezeichnung für Fette und fettähnliche Stoffe (Lipoide) mit nicht einheitlicher organisch-chemischer Struktur. Sie sind wasserunlöslich, aber in organischen Lösungsmitteln wie Benzin, Benzol, Ether, Chloroform, Methanol oder Aceton löslich.
Lipide sind ein wichtiger Bestandteil der Nahrung.
- 1 Gramm Fett enthält dabei 39 kJ = 9 kcal an Energie. 1 Gramm Zucker nur 4 kcal.
- Lipide sind also eine sehr energiereiche Nahrungsquelle
- Es gibt einige Lipide in der Nahrung, die der Mensch nicht selbst herstellen kann.
- Essentielle Lipide z. B.: Linol- und Linolensäure
- Lipide als Triglyzeride sind ein wichtiger Speicherstoff für Energie (Zucker dagegen werden in viel geringerer Menge in Form von Glykogen in der Leber gespeichert.)
- Lipide in der Haut schützen vor Verletzungen, da sie eine Polsterfunktion haben. Auch wichtige Organe können durch einen Fettmantel geschützt werden. Ebenso sind sie ein guter Kälteschutz in der Haut
- Lipide sind ein wichtiger Teil der Zellmembranen. (oft abgewandelte Lidide oder Verbundlipide wie Cholesterin, Tocopherol (=Vitamin E), Phospholipide oder Glycolipide (=Lipide mit Zuckeranteil)
- Lipide erleichtern als Gallensäuren die Fettverdauung. (Emulgatorwirkung)
- Lipoproteine (= Fetteiweißverbindungen) erleichtern den Fetttransport im Blut, da sie kleine Kugeln um die völlig wasserunlöslichen Triglyzeride bilden.
- Fettsäuren sind spezielle organische Säuren (unverzweigte Monocarbonsäuren), die aus einer - COOH Gruppe (=Carboxylgruppe) und einer unterschiedlich langen Kohlenwasserstoffkette (=Rest, z.B. R1)bestehen.
- Natürliche Fettsäuren bestehen in der Regel aus einer geraden Zahl von Kohlenstoffatomen und sind unverzweigt. Fettsäuren unterscheiden sich durch die Anzahl der C-Atome (Kettenlänge) und die Anzahl und Position ihrer Doppelbindungen. Fettsäuren ohne Doppelbindungen nennt man gesättigt. Ungesättigte Fettsäuren besitzen eine oder mehrere Doppelbindungen zwischen den Kohlenstoffatomen der Kette. Essentielle Fettsäuren sind solche, die ein Organismus nicht aus anderer Nahrung synthetisieren kann.
- Die Natrium- oder Kalium-Salze der höheren Fettsäuren sind die Seifen.
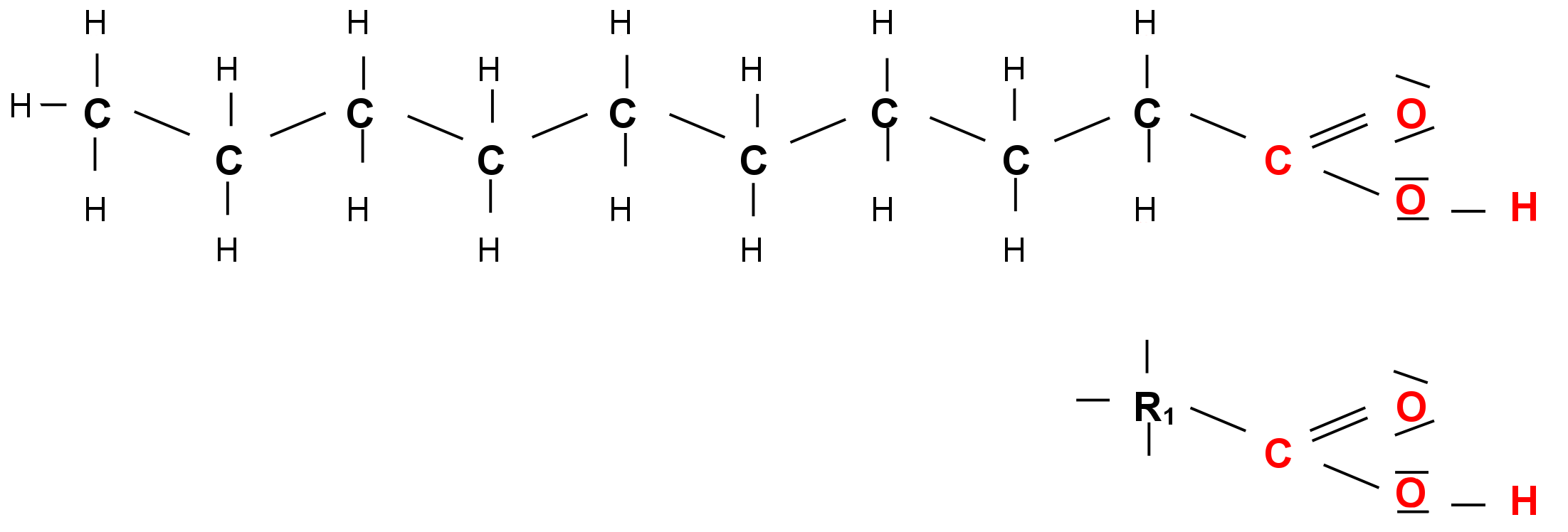
Fettsäuremolekül und Kurzschreibweise
Wiederholungsfragen Cytologie und Zellorganellen und Zellaufbau:
Membranen und Membranvorgänge
- Wie sind Membranen aufgebaut. Erstelle ein Modell.
- Welche Art von Proteinen gibt es im Zusammenhang mit Membranen?
- Welche Aufgaben haben sie?
- In der Mittelstufe hast Du vielleicht Chlorophyll aus Blättern extrahiert. Dazu hast Du die Blätter erst gekocht und dann in Spiritus gelegt. Erkläre, warum beide Schritt notwendig sind.
- Was sind Oberflächenproteine, welche Aufgaben haben sie?
- Erstelle eine Übersicht über Möglichkeiten für Feststoffe und Flüssigkeiten in Zellen und hinaus zu gelangen. Was sind in diesem Zusammenhang „Translokatoren“? Was ist in disem Zusammenhang „Endocytose“ / „Exocytose“?
- Was ist Plasmolyse (Deplasmolyse). Erkläre genau mit einer guten Skizze und einer Beschreibung aller Vorgänge?
- Was passiert mit Teer in der Lunge nach einiger Zeit? Wo bleibt er bei starken Rauchern?
- Warum sind gegossene Pflanzen praller und straffer als „durstige“?
- Wie kann es zu Organabstoßung nach einer Transplantation kommen?
- Zur Beschreibung von Blutgruppen teilt man diese in das AB0-System ein (A, B, AB und 0). Hinter den Buchstaben verbergen sich 2 verschiedene Oberflächenproteine der roten Blutkörperchen.
a) Erkläre, warum es mit nur zwei verschiedenen Oberflächenproteinen vier Blutgruppen geben kann (zur Not: https://de.wikipedia.org/wiki/Blutgruppen)
b) Weiße Blutkörperchen verklumpen in der Immunabwehrreaktion körperfremde Proteine. Erkläre, wie es zu tödlichen Blutinfusionen kommen kann? (Mit anderen Worten: Wieso muss man auf die Blutgruppe beim Spenden achten?)
c) Welche Blutgruppen sind für jemanden geeignet, der Blutgruppe A hat? (oder AB?)
d) Was versteht man unter Universalspenderblut und Universalempfänger? - Starker koffeinhaltiger Tee (wie auch Kaffee) (v.a. mit Zucker) gilt bei Ernährungsbiologen nicht als „Flüssigkeitsspender“ für den Körper. Beide führen auch zu vermehrtem Harndrang (v.a. im Vergleich mit der gleichen Menge Wasser). Erkläre warum Ernährungsbiologen dieser Meinung sind.
- Wenn man reife, feste Erdbeeren stark zuckert, verlieren sie Saft und nach einem Tag sind sie sehr weich. Erkläre die Zusammenhänge.
- Welche Wirkungen hat Nikotin, welche hat Zigarettenteer auf den menschlichen Körper?
- Erkläre, auf welchem Weg Giftstoffe, wie z.B. Schwermetalle in den Körper eindringen.
Versuch an Amöben:
a) Eine Amöbe wird in der Mitte in zwei Hälften zerteilt. (Das zellkernhaltige Stück lebt weiter). Nach einigen Tagen stirbt das zweite ab. Erkläre
b) Pflanzt man dem kernlosen Stück den Kern einer anderen Amöbe ein, lebt das Fragment weiter und entwickelt sich normal. Erkläre
- Freies Lehrbuch Biologie: 06.05 Gewebe
- Freies Lehrbuch Biologie: 07.01 Bakterien und Milchprodukte
- Freies Lehrbuch Biologie: 07.02 Infektionen und Immunabwehr - Feinde des Körpers
- Freies Lehrbuch Biologie: 07.03 Infektionen und Immunabwehr - Das Immunsystem und seine Abwehrreaktionen
- Freies Lehrbuch Biologie: 07.04 Infektionen und Immunabwehr - Allergien
- Freies Lehrbuch Biologie: 07.05 Infektionen und Immunabwehr - HIV und AIDS
- Freies Lehrbuch Biologie: 08.01 Zelluläre Grundlagen der Vererbung
- Freies Lehrbuch Biologie: 08.02 Zellzyklus, Mitose und Meiose als genetische Grundlagen
- Freies Lehrbuch Biologie: 08.03 Mendel und die klassische Genetik I
- Freies Lehrbuch Biologie: 08.04 Das Hormonsystem des Menschen
- Freies Lehrbuch Biologie: 08.05 Geschlechtsorgane, Geschlechtshormone und der weibliche Zyklus
- Freies Lehrbuch Biologie: 08.06 Entwicklungsbiologie I - Schwangerschaft und Verhütung
- Freies Lehrbuch Biologie: 08.07 Entwicklungsbiologie II - Wachsen und Erwachsenwerden
- Freies Lehrbuch Biologie: 08.08 Geschlechtskrankheiten
- Freies Lehrbuch Biologie: 08.10 Blut und Blutgruppen
- Freies Lehrbuch Biologie: 08.09 Erbkrankheiten I
- Freies Lehrbuch Biologie: 08.11 Entwicklungsbiologie III
- Freies Lehrbuch Biologie: 08.12 Molekulare Grundlagen der Vererbung (Molekulargenetik)
- Freies Lehrbuch Biologie: 08.13 Aminosäuren, Eiweiße, Enzyme und die Biokatalyse
- Freies Lehrbuch Biologie: 08.14 Variabilität des Erbguts, Erbkrankheiten und Stammbaumanalyse
- Freies Lehrbuch Biologie: 08.15 Klassische Genetik II
- Freies Lehrbuch Biologie: 08.16 Erbkrankheiten II
- Freies Lehrbuch Biologie: 08.17 Gentechnik
- Freies Lehrbuch Biologie: 09.02 Zigaretten und die Nikotinsucht
- Freies Lehrbuch Biologie: 10.06 Vitamine und Mineralsalze
- Freies Lehrbuch Biologie: 10.08 Zellatmung - Dissimilation
- Freies Lehrbuch Biologie: 11.01 Grundlagen und Begriffe der Verhaltensforschung
- Freies Lehrbuch Biologie: 11.02 Klassische Ethologie und angeborenes Verhalten
- Freies Lehrbuch Biologie: 11.03 Erworbene und erlernte Verhaltensanteile
- Freies Lehrbuch Biologie: 11.04 Sozialverhalten und Verhaltensökologie
- Freies Lehrbuch Biologie: 12.01 Evolution der Tiere und Pflanzen als Ursache der Vielfalt der Lebewesen
- Freies Lehrbuch Biologie: 12.02 Chemische Evolution - wie das erste Leben entstand
- Freies Lehrbuch Biologie: 12.03 Evolutionstheorien
- Freies Lehrbuch Biologie: 12.04 Unsere Vewandten - Affen