Kapitel 12.01: Evolution der Tiere und Pflanzen als Ursache der Vielfalt der Lebewesen

Der Schwanz eines Pfaus ist ein Beispiel für sexuelle Selektion
Ernst Mayr: „Evolution ist der wichtigste Begriff in der gesamten Biologie“.
Säugetiere sind Wirbeltiere
Lust auf ein Spiel aus Klasse 5? Genau, wir fangen nochmal ganz von vorne an. ;-)
Ordne die Dreiecke so an, dass zueinander passende Begriffe sich berühren. Eine Linie stellt eine Außenseite dar.

Die Lösung ist auf der letzten Seite. :-)
Grundfragen und Aufgaben der Evolutionsforschung
- Wie ist das Leben entstanden?
- Gab es einen Schöpfer? Gab es eine zufällige „Urzeugung“?
- Wie reagieren anorganischen Stoffe zu organischen?
- Welche genetischen Grundlagen sind für eine Entwicklung notwendig?
- Wie ist ein sich selbst reproduzierendes Protein-DNA-System entstanden, welches die Basis aller Lebensvorgänge bildet?
- Welche Fakten sprechen für eine gemeinsame Abstammung aller Lebewesen?
- Auf welchen Mechanismen beruhen anatomische, physiologische, ethologische Gemeinsamkeiten von Lebewesen?
- Wie lassen sich Fossilfunde einordnen und so für die Stammbaumrekonstruktion nutzen?
- In welchen Phasen/ Stufen ist die Evolution abgelaufen?
- Inwiefern unterscheiden oder ähneln sich rezente (heute lebende) und fossile Arten?
- Welche Evolutionsfaktoren gibt es?
Die Entwicklung der Wirbeltiere - Abstammung und Verwandtschaft

Du kennst sicher einige Haustiere wie den Hund, die Katze oder das Hausschwein. Diese stammen von Wildtieren ab und wurden vom Menschen bewusst in vielen Jahrhunderten gezüchtet. So wurde das Hausschwein aus dem Wildschwein gezüchtet und der Hund stammt vom Wolf ab.
Daraus folgt also, dass sich die Arten über viele Generationen hinweg verändern.
Also kann man vermuten, dass auch die verschiedenen Wirbeltierklassen voneinander abstammen. Es dauerte aber Jahrmillionen, bis sich die heutige Vielfalt bildete.
Von der Natur werden dabei Eigenschaften, wie z.B. das Haarkleid, in der Regel nur einmal „erfunden“, diese neuen Eigenschaften werden dann im Stammbaum weitervererbt, umgewandelt oder sie verschwinden völlig.
Historische Betrachtung der Evolution und der Entwicklung des Evolutionsgedankens
Biologen schätzen, dass auf der Erde heute etwa 1,5 Millionen verschiedene Tierarten und über
400 000 verschiedene Pflanzenarten leben. Jeder Lebensraum wurde von ihnen erschlossen. Diese Artenvielfalt ist erstaunlich und hat schon im Altertum die Menschen beschäftigt.
Der Frage nach der Herkunft dieser Vielfalt an Lebewesen (Woher und warum?) galt das Interesse der Menschheit seit Anbeginn.
a) Die Antike
Aristoteles (385-322 v. Chr.) war Begründer der Naturphilosophie, welche durch christliche Strömungen im Mittelalter mit dem Schöpfungsbericht in Einklang gebracht worden. Diese Schöpfungsdarstellung, die auch heute noch in jeder Bibel zu finden ist, galt bis ins 18. Jahrhundert als unantastbare Wahrheit! Jeder, der dieser widersprach, riskierte, als Ketzer verbrannt zu werden.
Kerngedanke dieser Schöpfungsphilosophie ist, dass die heutige Vielfalt durch einen einmaligen Schöpfungsakt entstand. (Allerdings ist die Bibel nicht immer wortwörtlich zu verstehen, selbst Augustinus warnte vor wörtlicher Auslegung des Schöpfungsberichtes!)
Nach Aristoteles kann man die Natur in Pflanzen und Tiere unterteilen. Die Tiere unterteilte er zudem in Bluttiere (Wirbeltiere) und blutlose Tiere (wirbellose Tiere). Durch genaue Beobachtungen konnte er damals schon die Delphine und Wale den Säugetieren zuordnen und er sah bei den Affen ein Bindeglied zwischen Mensch und Tier.
Aristoteles (so wie auch Platon 427 - 347 v. Chr.) glaubte an eine dualistische Natur:
a) die stoffliche Welt der Körper und Materie.
b) die unstoffliche Welt der Seelen, des Geistes, der Ideen sowie der Lebenskraft, welche er „vis vitalis“ nannte.
Er glaubte zum Beispiel auch, und das ist ihm nicht vorzuwerfen, wenn man bedenkt, wann diese Ideen entstanden sind und welche Mittel damals nur zur Verfügung standen, dass Lebewesen durch spontane Urzeugung aus Schlamm entstehen (vor allem Fische, Würmer und Insekten).
In vielen philosophischen Vorstellungen und Religionen hat sich diese Vorstellung bis heute gehalten.
b) Schöpfungsgeschichte:
Die Schöpfungsgeschichte der Bibel ist nicht die einzige. Viele Kulturen glauben, dass die frühe Menschheit durch Geister, Götter und Dämonen geschaffen oder beeinflusst wurde. Je nach Kultur muss man diese Götter mehr oder weniger milde stimmen oder besänftigen, um Katastrophen, wie Stürme, Epidemien oder Brände zu vermeiden. Dazu dienten kultische Rituale.
Für den europäischen Kulturkreis ist die jüdische Schöpfungsgeschichte die bedeutendste. Sie entstammt dem Alten Testament und wurde vermutlich 1000 – 500 v. Chr. aufgezeichnet. Der wesentliche Gedanke ist das gleichzeitige Entstehen allen Lebens als Produkte eines Schöpfungsaktes. Die Arten seien unveränderlich.
1. Buch Mose - Genesis, Altes Testament:
„Dann sprach Gott: Es lasse
sprießen die Erde grünende,
samenhaltende Kräuter und
fruchttragende Bäume, die
Früchte bringen nach ihrer
Art, Früchte, die in sich
Selbst ihren Samen tragen
auf Erden.“
„So schuf Gott die großen
Seeungeheuer und all die
lebenden Wesen, die sich
regen, von denen das Wasser
wimmelt, nach ihren Arten,
und alle beschwingten
Vögel in ihren Arten.“
„So machte Gott das Wild
des Feldes nach seiner Art,
das Vieh nach seiner Art
und alles Gewürm, das auf
dem Boden kriecht,
nach seiner Art.“
„Dann sprach Gott:
Lasst uns den Menschen machen
als unser Ebenbild, uns ähnlich.“
„Da bildete Gott der Herr
den Menschen aus dem Staub
der Erde und hauchte ihm den
Odem des Lebens ins Angesicht.“
Schöpfungsgeschichte: => Arten galten als etwas unveränderliches.
c) Das Mittelalter
Viele Forscher des Mittelalters sahen auch keinen Grund, die herrschende Philosophie der Schöpfung anzuzweifeln. Paracelsius (1498-1541) war beispielsweise bemüht, künstliches Leben auf diesem Wege herzustellen. Er nannte das zu erschaffende Wesen „Homunculus“.
Der Erbauer des Mikroskops, der Holländer Antoni van Leeuwenhoek, entdeckte 1675 unter seinem Mikroskop als Erster Bakterien und Einzeller. Er hielt an der „Schlammzeugungstherorie“ fest.
Selbst Newton, Descartes und Immanuel Kant hielten an der Theorie der Schöpfung fest!
Wenigen Wissenschaftlern und Philosophen kamen Zweifel. Zu den wenigen gehörten der engl. Arzt Harvey (1578-1657) und der italienische Arzt Redi, welcher durch einen Versuch 1650 zeigte, dass aus faulem Fleisch unter einem Fliegenschutz keine Würmer wachsen.
Louis Pasteur bewies erst 1860, dass die Mikroorganismen in faulem Fleisch von außen hinein gelangt sein müssen. Damit war die Theorie von der Urzeugung widerlegt. An der Schöpfung der Lebewesen in einem einmaligen Schöpfungsakt wurde dennoch festgehalten.
d) Carl von Linné (1707-1778) und die Lehre von der Konstanz der Arten
Linné war der erste Biologe, der eine umfassende Systematisierung der Organismen nach anatomischen und physiologischen Merkmalen durchführte. Eine Veränderbarkeit der Arten erkannte er auch nicht (=> binäre Nomenklatur).
taxonomische Kategorie = Taxa
Der schwedische Biologe Carl von Linné (1707-1778) stellte 1735 in seinem Buch, dem „Systema Naturae“ ein von ihm entwickeltes Ordnungssystem der Tier- und Pflanzenwelt vor. Dieses galt bis in die 1980er Jahre als einzige Möglichkeit, Tiere und Pflanzen zu klassifizieren. Linnes Nomenklatur beruhte allein auf äußerlichen Merkmalen der Tiere und Pflanzen. Seine Leistung ist aber enorm, da er sehr sauber sehr viele Tiere (4230) und Pflanzen (14000) kategorisierte und erstmals Verwandtschaften zwischen Arten feststellte. Verwandte Arten ordnete er in Gattungen ein.
Viele dieser Verwandtschaften sind heute durch genetische Untersuchungen bestätigt worden. Allerdings zeigen heutige Untersuchungen auch Unterschiede, so stellt die damalige Gruppe der Reptilien keine einheitliche Gruppe dar. Schildkröte, Krokodile und Echsen sind gar nicht so nah miteinander verwandt.
Linné ging ebenfalls von einer einmaligen Schöpfung der Lebewesen aus und glaubte demzufolge ebenfalls, so wie alle vor ihm auch, an die Artkonstanz, die Unveränderbarkeit der Lebewesen.
Das Vorhandensein ausgestorbener Arten, bekannt durch fossile Funde, wurde z.B. mit der biblischen Sintflut, in welcher viele Arten gestorben sein sollen, erklärt.
Aber Linné war auch ein schlauer Fuchs, um den Menschen (als Gattung „Homo“) aufgrund seiner Verwandtschaft zu den Menschenaffen in seinen Stammbaum der Tiere aufnehmen zu können, was ja der Lehre der Kirche widersprach, nannte er diese Gruppe „Primaten“ und bezeichnete den Menschen als Krönung der Schöpfung. Er postulierte das als biblischen Schöpfungsplan, welche er nun offengelegt habe. So konnte die Kirche nicht widersprechen. :-)
Das System der Lebewesen nach Carl von Linné
|
Systematische Kategorie |
Einordnung „Wolf“ |
|
Reich |
Tierreich |
|
Unterreich |
Vielzeller |
|
Stamm |
Wirbeltiere (Vertebrata) |
|
Klasse |
Säugetiere (Mammalia) |
|
(Unterklasse) |
Höhere Säugetiere (Eutheria) |
|
Ordnung |
Raubtiere (Carnivora) |
|
Familie |
Hunde (Canidae) |
|
Gattung |
Canis |
|
Art |
Wolf (Canis lupus) |
Merksatz zur Reihenfolge:
Klasse Ordnung herrscht in der Familie, wenn der Gatte artig ist.
Problematisch sind nach diesem System die Einordnungen der Fossilien. Wie kann man z.B. frühe Formen der Gattung „Homo“ (Mensch) einordnen, wenn sie sich in nur wenigen Merkmalen unterscheiden?
Dieses System gilt heute als veraltet! Man findet es aber noch in vielerlei Büchern - also bitte nicht verwirren lassen.
e) Cuviers Katastrophentheorie
Georges Léopold Chrétien Frédéric Dagobert, Baron de Cuvier (oder auch Georges Cuvier für seine Freunde *g*) (1769-1832), erklärte die Absenz vieler Arten durch eine hypothetische „Katastrophentheorie“, welche zum plötzlichen Aussterben vieler Lebewesen führte.
=> Die überlebenden Arten führten eine Neubesiedlung durch.
Aber diese Theorie ließ vieles unberücksichtigt. Die abgestufte Ähnlichkeit von Arten und die bei vielen Arten „fließenden“ Artgrenzen konnten so nicht erklärt werden.
f) Die Abstammungstheorien von Lamarck (1744-1829) und Charles Darwin (1809-1882)
Details zu den jeweiligen Theorien siehe Kapitel 12.03.
Beide erkannten, dass Arten nicht konstant sind. Stattdessen findet eine stetige Entwicklung der Lebewesen statt.
=> Evolution
Die Evolution beschreibt den Vorgang der Entwicklung der Lebewesen von einfachen zu komplexen Formen. Evolution ist ebenfalls die Erklärung dafür, dass alle Lebewesen durch gemeinsame Abstammung miteinander verwandt sind und vermutlich von einem gemeinsamen ersten Lebewesen abstammen.
Übersicht der Theorien
Linné: Lehre von der Konstanz der Arten (1737)
„Alle heutigen Arten sind durch einen Schöpfungsakt entstanden“.
Lamarck: Abstammungstheorie (Descendenztheorie) (1809)
„Lebewesen entwickeln sich von einfachen zu komplexen Lebewesen. Entscheidend für diese Entwicklung ist der Gebrauch oder Nichtgebrauch von Organen und die folgende Weitervererbung von so erworbenen Veränderungen.“
(Theorie von der „Wandelbarkeit der Arten“)
Cuvier: Katastrophentheorie (1828)
„Fossile Funde wie Versteinerungen von nicht mehr existierenden Arten sind Reste von riesigen Naturkatastrophen (z.B. Sintflut)“.
Cuvier war entschiedener Gegner der Abstammungstheorie!
Darwin: Neue Version der Abstammungstheorie (1859)
„The Origin of Species by Means of Natural Selection!“
Zwischen den Individuen einer Art bestehen Unterschiede. Diese sind zufällig. Die Natur selektiert von diesen Individuen die geeignetsten aus: „Survival of the fittest“.
Mendel: Vererbungsgesetze (1865)
Die Mendelschen Regeln beschreiben die Vielfalt der Lebewesen. Eine Evolution der Arten erkannte Mendel jedoch nicht
Haeckel: Biogenetisches Grundgesetz (1866-1969)
„Die Ontogenese ist eine verkürzte Wiederholung der Phylogenese!“
Ernst Haeckel ist der bedeutendste deutsche Vertreter der Abstammungslehre.
Zusatzinformationen:
http://de.wikipedia.org/wiki/1._Buch_Mose
http://www.bibel-online.net/geschichte.html (Zeittafel der Bibelentstehung)
Was bedeutet nun Evolution?
Heute weiß man, vor allem durch die Pionierarbeit Charles Darwins, dass alle heutigen Tier- und Pflanzenarten sich in einem permanenten Entwicklungsprozess befinden. Diese Arten haben sich durch diese Entwicklung im Laufe sehr großer Zeiträume (Jahrmillionen), aus einfacheren Lebewesen zu komplexeren entwickelt. Am Anfang stand vermutlich nur eine Art, ein sehr einfacher Einzeller.
So sind im Grunde alle Lebewesen miteinander verwandt, wobei die Verwandtschaft zu einigen Lebewesen geringer ist als zu anderen. ;-)
Dabei liegt der Evolution das Prinzip zugrunde, dass die Entwicklung immer von einfacheren Formen und Strukturen zu komplexeren abläuft!
Achtung Begriffstrennung: Die Evolution ist ein Vorgang und keine Theorie! Das die Evolution stattfindet, ist unumstritten, es gibt Millionen von Belegen!
Die Erklärung, wie sie stattfindet, wird in Darwins Evolutionstheorie oder auch der „Synthetischen Evolutionstheorie“ erklärt. Dabei handelt es sich aber tatsächlich um Theorien, da niemand genau sagen kann, wie das komplexe System des Lebens auf diesem Planeten funktioniert.
Der Artbegriff
Die kleinste Einheit der Evolutionsforschung ist die Art. Nun ist es nicht immer einfach, zu bestimmen, welche Tiere zur gleichen Art gehören (oder evtl. doch nur zu verschiedenen Rassen). Linné definierte als Erster genau den Begriff der Art (=Spezies). Er tat dies nach morphologischen Merkmalen, also äußerlichen Merkmalen der Gestalt und des Körpers:
Linné: Eine Art ist eine Gruppe von Lebewesen, die in ihrem inneren Aufbau und ihrer
äußeren Gestalt übereinstimmen.
Diese Definition gilt heute als überholt. Es gibt heute mehrere Definitionen. Gut zu gebrauchende Definitionen sind die biologischen Definitionen des Artbegriffs:
a) Biologischer (= genetischer) Artbegriff:
Diese Definition ist die biologisch eindeutigste. Es gibt verschiedene Formulierungen:
- Eine Art umfasst alle Individuen, die sich miteinander auf natürliche Weise faktisch und potenziell fortpflanzen können und fruchtbare Nachkommen hervorbringen.
- Alle Mitglieder einer Art können miteinander fruchtbare (!) Nachkommen zeugen. Unter Biologen oft etwas salopp gesagt: Alles, was sich scharrt und paart, gehört zu einer Art. ;-)
- Ernst Mayr (1969): „Arten sind Gruppen von sich miteinander kreuzenden natürlichen Populationen, die von anderen Gruppen reproduktiv isoliert sind.“
- Phylogenetisches (evolutionäres) Artkonzept: Als Art wird eine monophyletische Abstammungsgemeinschaft definiert, welche (aus einer bis vielen Populationen) innerhalb einer bestimmten Zeitspanne lebt (gekennzeichnet durch eine Linie im Stammbaum). Eine Art beginnt nach einer Artaufspaltung (Verzweigung im Stammbaum) und endet, wenn die neue Linie sich erneut spaltet oder alle Mitglieder dieser Art aussterben.
- Populationsgenetisches Artkonzept: Eine Art besteht aus mindestens einer Population, deren Genpool gegen Populationen anderer Arten generativ isoliert ist.
b) Morphologischer Artbegriff:
Zu einer Art gehören alle Organismen, die in ihren wesentlichen Merkmalen oder im Verhalten übereinstimmen (z.B. Aussehen, Anatomie, Verhalten).
c) Ökologischer Artbegriff:
Tiere, die sich eine ökologische Nische teilen, gehören zu einer Art (diese Definition ist kaum zu gebrauchen, weil sie sehr ungenau ist!).
Zusatzinformationen:
http://de.wikipedia.org/wiki/Art_%28Biologie%29
Weitere wichtige Begriffe
Genpool (auch „Genschatz“ oder „Genfundus“)
Der Genpool ist die Gesamtheit der genetischen Informationen und aller Genvariationen (Allele), über die eine Art verfügt. Jedes Individuum einer Art hat aber nur einen Bruchteil der insgesamt vorhandenen Allele.
http://de.wikipedia.org/wiki/Genpool
Population:
Die Gesamtheit der Individuen einer Art, die in einem bestimmten Verbreitungsgebiet leben und sich untereinander fortpflanzen (Fortpflanzungsgemeinschaft) nennt man Population. Eine Art besteht in der Regel aus vielen Populationen.
Rassen (=Unterart):
Eine Rasse ist eine Einheit unterhalb des Artbegriffes, also Gruppen von Individuen innerhalb einer Art, welche sich in mindestens einem reinerbigen Merkmal voneinander unterscheiden aber dennoch fortpflanzungsfähige Nachkommen mit den Individuen andere Gruppen haben können.
Kommen weitere Merkmale der Unterschiedlichkeit hinzu, kann eine Trennung und Auseinanderentwicklung zu zwei neuen Arten stattfinden. Phylogenetiker verwenden häufiger den moderneren Ausdruck Unterart oder Subspezies.
Zusatzinformationen:
http://de.wikipedia.org/wiki/Rass
Wichtige Persönlichkeiten der Evolutionsforschung
- Linne pd: http://de.wikipedia.org/wiki/Bild:Carolus_Linnaeus_%28cleaned_up_version%29.jpg
- Cuvier:pd http://de.wikipedia.org/wiki/Bild:Georges_Cuvier_large.jpg
- Lamark CC-By-SA: http://de.wikipedia.org/wiki/Lamarck; Author Valérie75; http://creativecommons.org/licenses/by-sa/3.0/deed.de
- http://commons.wikimedia.org/wiki/Jean-Baptiste_Lamarck
- Darwin: pd http://de.wikipedia.org/wiki/Bild:Charles_Darwin_by_Julia_Margaret_Cameron.jpg
- http://commons.wikimedia.org/wiki/Charles_Darwin
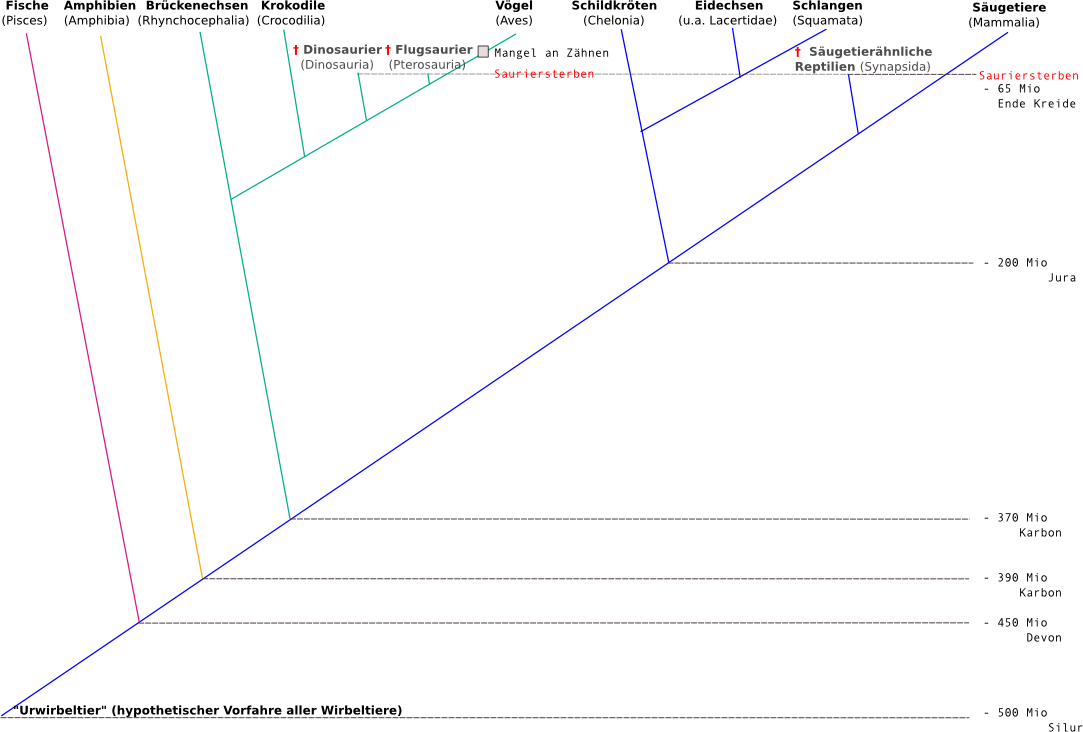
Der Stammbaum der Wirbeltiere (=Vertebrata)
Die Wirbeltiere sind miteinander verwandt, d.h. sie hatten gemeinsame Vorfahren. Einer dieser Vorfahren ist vermutlich das so genannte Urwirbeltier. Es unterschied sich von den Weichtieren und anderen damals lebenden Tierarten durch die Ausbildung einer sehr ursprünglichen Wirbelsäule.
Es lebte vermutlich vor vielen hundert Millionen Jahren als Meeresbewohner und kann als Verbindung zwischen wirbellosen Tieren und Wirbeltieren gesehen werden. Von ihm stammen die ersten Fische ab. In den folgenden Millionen von Jahren haben sich daraus die Amphibien, die Reptilien, die Vögel und die Säugetiere entwickelt. Heute kennt man ca. 55.000 Wirbeltierarten.
Genetische Untersuchungen konnten dies aber nicht komplett bestätigen:
- Die klassischen „Reptilien“ sind keine einheitliche Gruppe (=Monophyllum), sondern ein Paraphylum und somit in zwei große Gruppen aufzuteilen.
- Die klassischen Fische sind vermutlich ebenfalls ein Paraphylum und aufzuteilen.
Gemeinsamkeiten aller Wirbeltiere:
- Knocheninnenskelett (Wirbelskelett als Achse)
- 2 Paar Extremitäten
- Gehirn (geschützt von Schädelkapsel)
- Rückenmark
- geschlossenes Blutkreislaufsystem
Die Gemeinsamkeiten lassen auf gemeinsame Vorfahren und eine Verwandtschaft der Tierarten schließen.
=> Die Entwicklung führt vom Einfachen zum Komplexen.
Zusatzinformationen:
Federn wurden auch bei manchen Arten der Dinosaurier gefunden. Daher nimmt man an, dass auch sie zumindest ansatzweise gleichwarm waren. Dies ist ein direkter Beweis für die Aufspaltung der klassischen Gruppe der Reptilien in zwei Gruppen!
http://de.wikipedia.org/wiki/Wirbeltiere
Aufgaben: Vererbung von Merkmalen
1. Ordne in der Gruppe die folgenden Begriffe in den Stammbaum ein:
Merkmale:
- Kiemen,
- Lunge,
- Eier mit Kalkschale,
- innere Befruchtung durch Penis, äußere Befruchtung,
- wechselwarm,
- gleichwarm,
- lebend gebärend (mit Gebärmutter!),
- Federn,
- Haare,
- Milchdrüse,
- kernlose rote Blutkörperchen,
- Knochen aus Knorpel und Kalk,
- Wirbelsäule,
- Haut ohne Federn Haare oder Schuppen,
- Seitenlinienorgan
- Hornschuppen,
- Zähne,
- 2 Beinpaare (mit Oberarm/-schenkel, Elle & Speiche/Schienbein & Wadenbein sowie die Hand-/Fußknochen und Fingern bzw. Zehen)
- Luftröhre,
- Augenlider,
- feuchte & stark luftdurchlässige Haut (ohne Federn, Haare, Schuppen)
- Krallen oder Nägel aus Horn,
- Harnblase,
- beschalte (aber noch recht weiche) Eier,
- 3 Gehörknöchelchen
- Extremitäten lang und schlank,
- lang gestreckte Halswirbel,
- aufrechter Gang auf den Hinterbeinen,
- Mangel an Zähnen
2. „Eine Linie beschreibt eine Art“! Ist diese Aussage richtig?

Was ist phylogenetische Forschung?
Das Wort Phylogenese (Stammesentwicklung) leitet sich vom griechischen Wort „philon“ (Stamm, Geschlecht) dem Wort „genessi“ (Geburt, Entstehung) ab. Es beschreibt unter anderem eine modernere und wissenschaftlichere Einordnung von Lebewesen in Stammbäume, um die stammesgeschichtliche Entwicklung und die biologische Evolution der Lebewesen nachzuvollziehen und zu verstehen. Das Prinzip des Einordnens von Lebewesen in phylogenetische Stammbäume nennt man auch Kladistik.
Einer der wichtigsten Begründer des phylogenetischen Forschungszweiges innerhalb der biologischen Disziplinen war Willi Hennig. Bevor er nach vererbbaren Merkmalen Stammbäume aufstellte, galt das klassische System von Carl von Linné (mit Dir sicher bekannten Begriffe wie Ordnung, Gattung, Familie usw.), welches Verwandtschaft vor allem nach äußerlichen Faktoren definierte (siehe auch den Abschnitt zu Linné weiter vorn). Die heutigen genauen genetischen Untersuchungsmethoden bestätigen Hennigs Arbeitsweise. Je genauer mittlerweile Gensequenzen analysiert werden, desto genauer werden die Stammbäume.
Die Konsequenz ist, das einige bekannte Einordnungen, die viele Jahre lang als richtig galten, neu getroffen werden mussten. So gibt es z.B. nicht die einheitliche Gruppe der Reptilien. Die Reptilien umfassen vielmehr drei Gruppen, mit optisch ähnlichen Kriechtieren, welche direkt miteinander verwandt und somit nicht einheitlich sind!
Der Vorteil des phylogenetischen Systems liegt in seiner Sichtweise auf vererbbare Merkmale (hinter denen sich ja Gene verbergen) und der Entwicklung der natürlichen Verwandtschaftsgruppen. Schwierig hingegen ist natürlich, dass die Entwicklung der Arten nicht beobachtet werden kann. Fossile Funde und deren Analyse stellen deshalb eine wichtige Grundlage der phylogenetischen Forschung dar.
Zum Erstellen von Stammbäumen sind u.a. folgende Arbeitsweisen möglich:
- Auswertung von äußeren und physiologischen Merkmalen (z.B. des Stoffwechsels) von lebenden Tieren und Fossilien.
- Vergleich der äußeren und physiologischen Merkmale von Lebewesen
- Analyse der Ontogenese (da sich in dieser die Stammesentwicklung widerspiegelt (siehe E.Haeckel))
- Analyse der DNA
Begriffe der Pyhlogenetik nach Hennig
a) Taxon
Eine Art wird bei Hennig Taxon genannt. Dabei wird nicht wie in der klassischen Stammbaumforschung nur von einer Art, wie z.B. dem „Hund“, ausgegangen. Eine Taxon ist grundsätzlich eine Linie im Stammbaum, welche mehrere Arten zusammenfassen kann (wurde in der klassischen Stammbaumforschung dann als Gattung, Ordnung usw. bezeichnet).
Taxa und deren Verwandtschaft werden in der Phylogenetik (auch Kladistik genannt) niemals nur anhand des Aussehens zusammengefasst. Grundlage ist immer das Vorhandensein von einheitlichen Genen! Diese sind bei genauer Betrachtung aber mit recht hoher Wahrscheinlichkeit auch optisch anhand des jeweilig ausgeprägten Merkmals erkennbar.
b) Autapomorphie, Plesiomorphie:
Der zentrale Begriff der Phylogenetik ist der der Autapomorphie. Er bezeichnet ein (aut)apomorphes Merkmal, welches als neu entwickeltes Merkmal eine Art von einer anderen verwandten Art unterscheidet. Die Autapomorphie stellt sozusagen eine „Neuerfindung der Natur“ dar (=evolutive Neuheit). Sie wird auch oft als abgeleiteter Merkmalszustand bezeichnet.
Demgegenüber gibt es auch die Plesiomorphie. Sie ist ein bereits vorhandenes, ursprüngliches Merkmal oder dessen Merkmalsausprägung. Plesiomorphe Merkmale sind bereits vor der jeweils betrachteten Stammeslinie entstanden und haben sich nicht im Laufe der Zeit verändert. Plesiomorphien sind also bei mehreren Arten zu finden.
Beispiele für Plesiomorphien:
Bei Menschen wäre dies z.B. das Vorhandensein von Eckzähnen, welche schon bei älteren Säugetierarten zu finden sind.
Ein weiteres Beispiel wäre das Merkmal „Vierfüßigkeit“ bei den landlebenden Wirbeltieren (Tetrapoden). Da alle Vertreter dieser Teilgruppe vier Gliedmaßen haben, können Verwandtschaftsverhältnisse mit diesem plesiomorphen Merkmal nicht bestimmt werden.
c) Synapomorphie und Symplesiomorphie:
Das eben genannte Merkmal „Vierfüßigkeit“ ist gleichzeitig eine Synapomorphie der Wirbeltiere, gegenüber der Gruppe der Fische, da die Vierfüßigkeit von den Tetrapoden neu erworben wurde. Der Begriff Synapomorphie (altgriechisch: Syn/ Sym = mit) ist ein untergeordneter Begriff. Er bezeichnet den gemeinsamen Besitz einer Apomorphie von mehreren sogenannten Schwestertaxa (=Schwesterarten) (z.B. Milchdrüsen bei allen Säugetieren).
Dementsprechend ist eine Symplesiomorphie ein ursprüngliches Merkmal, welches mehreren Taxa zugeordnet werden kann, aber keine Neuentwicklung darstellt! Sie ist also eine Übereinstimmung in plesiomorphen Merkmalszuständen.
Beispiele für Symplesiomorphien:
- die Wirbelsäule aller Säugetiere
- die oben genannte Vierfüßigkeit aller Tetrapoden
- der fünfstrahlige (pentadaktyle) Bau der Hände z.B. bei Eidechsen, Mäusen und Menschen
Unterscheide:
- Apomorphien sind Merkmale, welche durch eine Art neu entstanden sind. (z.B. Flügel als Autapomorphie der geflügelten Insekten, Deckflügel als Autapomorphie der Käfer)
- Synapomorphien begründen hingegen ein Schwestergruppenverhältnis. Eine Stammart hatte die Apomorphie „erfunden“ und diese dann an die nachfolgenden Arten vererbt. (z.B. zwei Flügelpaare sind eine Synapomorphie welche man bei Libellen und Heuschrecken findet).
=> Die Zusammenfassung von Taxa (Arten) aufgrund von Symplesiomorphien führt zu paraphyletischen Gruppen. (Gruppe, welche nicht die komplette Verwandtschaftslinie mit einschließt, also Schwestergruppen außen vor lässt).
=> Zusammenfassungen aufgrund von Synapomorphien führen zu monophyletischen Gruppen (Eine monophyletische Gruppe beinhaltet eine Stammart und alle Nachfahren dieser Stammart).
Merkmale, die bei diesen Vergleichen betrachtet werden, können übrigens anatomische, physiologische (=Stoffwechsel) sowie physiologische oder genetische Merkmale (gleiche DNA Sequenz) sein. Selbst der Verlust eines Merkmals kann als Autapomorphie gesehen werden, wenn er eine Weiterentwicklung darstellt (z.B. Wegfall der Schwanzwirbelsäule beim Menschen).
Plesiomorphe Merkmale sind also in der Regel die einfacheren und ursprünglicheren und apomorphe Merkmale sind die komplizierteren, höher entwickelten Merkmale.
Vergleiche: Monophylum und Paraphylum
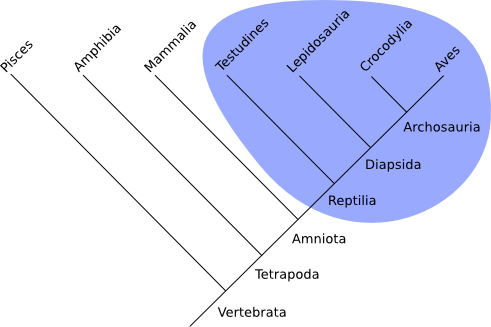
Monophylum (monophyletische Gruppe):
=> direkte Verwandtschaft!
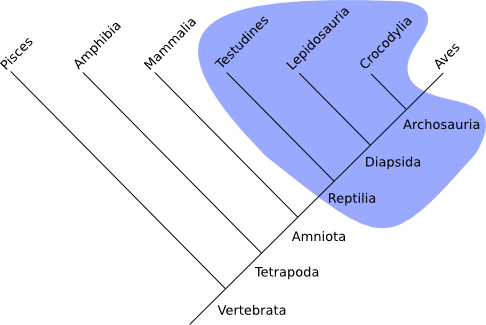
Paraphylum (paraphyletische Gruppe):
Verwandtschaft, aber nicht die komplette Gruppe in inkludiert! So ist die früher oft genannte Gruppe der „Reptilien" eine paraphyletische Gruppe, da sie die Vögel nicht mit einschließt.

Polyphylum (polyphyletische Gruppe):
Ist zu nichts zu gebrauchen *g*. Kann nicht als Beweis für direkte Verwandtschaft verwendet werden!
Quelle aller drei Bilder: GNU-Lizenz für freie Dokumentation sowie Creative Commons „Namensnennung-Weitergabe unter gleichen Bedingungen 3.0 Unported; http://de.wikipedia.org/wiki/GNU-Lizenz_für_freie_Dokumentation; http://creativecommons.org/licenses/by-sa/3.0/deed.de
1. Wikivommonsuser JacekFH, Thanks - http://commons.wikimedia.org/wiki/Image:Monophyletic.svg
2. Wikivommonsuser JacekFH, Thanks - http://commons.wikimedia.org/wiki/File:Paraphyletic.svg
2. Wikivommonsuser JacekFH & Zeimusu, Thanks; http://commons.wikimedia.org/wiki/Image:Polyphyletic-mammals-birds.svg
d) Wozu braucht man Autapomorphien?
Autapomorphien sind das zentrale Beweismittel bei der heutigen Stammbaumforschung! Ist ein Merkmal als Autapomorphie erkannt, so stellt es das Alleinstellungsmerkmal einer Art dar. Diese Art kann nun als Taxon und als Monophyla begründet werden. Wissenschaftlich bewiesen ist es aber erst, wenn zugleich eine plesiomorphe Merkmalsalternative gefunden ist! Als Autapomorphie wird ein Merkmal dann sicher erkannt und zugeordnet, wenn es innerhalb der eigenen Stammlinie (=Taxons) homolog vorhanden ist und bei allen Schwestergruppen nicht vorhanden ist.
Ein Beispiel zur Verdeutlichung:
Das Taxon der Insekten hat als begründende Autapomorphie zwei Flügelpaare. Damit ist das Insektentaxon Pterygota (Fluginsekten) begründet. Die Vorfahren der heutigen Fluginsekten waren flügellos. Diese Flügellosigkeit der Urinsekten (und übrigens auch aller übrigen Gliederfüßer wie Spinnen, Tausendfüßern, Krebsen usw.) ist also eine Plesiomorphie.
Betrachtet man jetzt die Pterygota (Fluginsekten) für sich, kann man das Merkmal „zwei Flügelpaare“ nicht mehr zur Rechtfertigung weiterer Arten verwenden. Für alle Arten innerhalb des Taxons Fluginsekten ist also das Merkmal „zwei Flügelpaare“ ein plesiomorphes Merkmal!
Erst deren weitere Veränderung, z.B. Umwandlung des vorderen Paares zu Flügelklappen (bei allen Käfern) stellt eine neue Autapomorphie dar, welche nun das Taxon der Käfer begründet.
Zusatzinformationen (vor allem, wenn die Begriffe noch verwirrend sind!):
http://de.wikipedia.org/wiki/Kladistik
Fische ein Monophylum?
Die Wirbeltiere stammen von den Fischen ab

Quelle Bild: cc-by-sa, Wikipedia.org; https://de.wikipedia.org/wiki/Kiefermäuler
f) Monophylum und Paraphylum
siehe: Kladistik
Zusatzinformationen:
- http://de.wikipedia.org/wiki/Kladistik
- http://de.wikipedia.org/wiki/Phylogenese
- http://de.wikipedia.org/wiki/Paraphylum#Verwandtschaftsverhältnisse
- http://de.wikipedia.org/wiki/Plesiomorphie
- http://de.wikipedia.org/wiki/Autapomorphie
- http://de.wikipedia.org/wiki/Willi_Hennig
- http://de.wikipedia.org/wiki/Systematik_%28Biologie%29
- http://de.wikipedia.org/wiki/Biogenetisches_Grundgesetz
- http://de.wikipedia.org/wiki/Ernst_Haeckel
- http://de.wikipedia.org/wiki/Homologie_(Biologie)
- http://de.wikipedia.org/wiki/Analogie_%28Biologie%29
Gliederung der Erdgeschichte
a) Archaikum und Proterozoikum
- Vor ca. 4,6 Milliarden Jahren ist die Erde entstanden.
- Aus der Zeit von vor ca. 3,5 Milliarden Jahren stammen die ältesten fossilen Funde. Eine genaue Datierung ist bei so großen Zeitabständen sehr schwierig.
- Vor ca. 2,5 Milliarden Jahren entstand erster freier Sauerstoff in der Atmosphäre.
- Vor 1,5 Milliarden Jahren entstanden die ersten Eukaryoten, also Zellen, welche über einen Zellkern verfügten.
- Vor ca. 700 Millionen Jahren sind erste Tierarten entstanden.
b) Paläozoikum (=Erdaltertum)
Kambrium: Zeitraum von -542 bis -500 Mio. Jahren
- bereits reiches Tier- und Pflanzenleben vorhanden, Leben kommt aber nur im Meer vor! Vermutlich stieg genau in diesem Zeitraum die Artenzahl enorm an. Man spricht von der „Kambrischen Explosion“.
- nachgewiesenes tierisches Leben: vermutlich alles wirbellose Gruppen (also Hohltiere, Trilobiten (Dreilappkrebse), Schnecken, Stachelhäuter sowie Gliederfüßer)
- nachgewiesenes pflanzliches Leben: Algen (Grün-, Rot- und Blaualgen)
- viele Versteinerungen erhalten
Ordovicium: Zeitraum von -500 bis -435 Mio. Jahren
- Kieferlose Urfische stellen die ersten nachgewiesenen Wirbeltiere dar.
- Algen entwickeln sich weiter und werden komplexer.
- Vermutlich sind kleine, gepanzerte, kieferlose, fischähnliche Tiere die ersten Wirbeltiere.
Silur: Zeitraum von -435 bis -400 Mio. Jahren
- Besiedlung des Festlandes durch Nacktfarne und bärlappartige Gefäßsporenpflanzen beginnt.
- Die Anzahl der wirbellosen Tierarten nimmt enorm zu.
- nachgewiesenes tierisches Leben im Meer: Korallen, Trilobiten, Kopffüßer, Stachelhäuter, Muscheln, Schnecken uvm.
- Entstehung der Panzerfische (Wirbeltiere!), welche bereits über einen Kiefer verfügen.
- Es gibt bereits drei Gruppen von Fischen (solche mit paarigen Brustflossen, Fische mit Hautknochen und Fische mit Kieferskelett (Vorfahren der heutigen Haie).
Devon: Zeitraum von -400 bis -350 Mio. Jahre
- Sumpflandschaften entstehen, starke Zunahme der Gefäßsporenpflanzen wie Schachtelhalme, Bärlappe, Urfarne usw.).
- Pflanzen haben schon echte Wurzeln und Wasserleitungssysteme im Inneren (Leitbündel und -gefäße).
- Blätter haben schon Spaltöffnungen.
- Im Meer starke Zunahme der Fische (Devon ist das Zeitalter der Fische), erstes Auftreten von Knochenfischen, Lungenfischen und Quastenflossern.
- Beginn der Landeroberung der Tiere => erste Gliederfüßer an Land (Insekten, Spinnen, Skorpione, Tausendfüßer sowie erste Amphibien), am Ende des Zeitalters vermutlich erste Landwirbeltiere (Ichthyostega).
Diese Eroberung des Festlandes durch Skorpione und Tausendfüßer war möglich, da sie bereits über einen Chitinpanzer als Verdunstungsschutz verfügten.
Karbon: Zeitraum von -350 bis -280 Mio. Jahren
- Farnwälder aus Bärlapp, Farnen und Schachtelhalmen entstehen (so genannte Steinkohlewälder, da sie die Grundlage der heutigen Steinkohle sind!). Typische Bäume sind Schuppenbäume, Siegelbäume, Riesenschachtelhalme.
- Samenfarne entstehen als Vorläufer der Nacktsamer.
- Gliederfüßer werden immer größer und nehmen zum Teil Riesenformen an (Urlibelle mit 80cm Spannweite!).
- Zu Beginn des Karbons sind Amphibien die dominierende Gruppe
- zum Ende des Karbons entstehen erste Reptilien.
Perm: Zeitraum von -280 bis -220 Mio. Jahren
- Baumfarne dominieren die Pflanzenwelt, erste Ginkogewächse bilden sich! Nacktsamer nehmen zu.
- Artensterben: Trilobiten und Panzerfische sterben aus.
- Reptilien beginnen die Dominanz, erste Saurier tauchen auf, frühe Säugetiere entstehen, spielen aber anzahlmäßig keine Rolle.
c) Mesozoikum (=Erdmittelalter):
Trias: Zeitraum von -220 bis -190 Mio. Jahren
- Buntsandstein, Muschelkalk und Keuper werden gebildet (Bodenformen).
- Erste Nadelbäume entstehen. Palmfarne dominieren aber anfangs.
- Große Amphibien (Riesenlurche) sind weit verbreitet.
- Reptilien v.a. die Anzahl der Saurier nimmt stark an Land, in der Luft und im Wasser zu
- erste Blütenpflanzen entstehen
- Vermutlich sind gegen Ende des Trias schon sehr einfache Säugetiere vorhanden.
Jura: Zeitraum von -190 bis -135 Mio. Jahren
- Zeitalter der Saurier (Hauptzeit der Saurier!), gegen Ende Entstehung des Archaeopteryx!
- Ginkobäume, Nadelbäume und Palmfarne dominieren.
- Ammoniten und Belemniten sind weit verbreitet.
- Erste Knorpelfische und erste Knochenfische entstehen.
- Erste Beuteltiere entstehen.
Kreide: Zeitraum von -135 bis -70 Mio. Jahren
- Erstes Auftreten von Bedecktsamern! Laubbäume und Gräser entstehen. Viele
Blütenpflanzen.
- Ammoniten, Belemniten sowie die Dinosaurier sterben aus.
- Knochenfische und Beuteltiere nehmen entsprechend in ihrer Anzahl zu!
- Erste hoch entwickelte Säugetiere! (Kloakentiere, Beuteltiere)
- Beginn des Reptiliensterbens
- Erste echte Vögel.
d) Känozoikum (=Erdneuzeit):
Tertiär: Zeitraum von -70 bis -2 Mio. Jahren (umfasst Paläozän, Eozän, Oligozän, Miozän, Pliozän)
- Blütenpflanzen beginnen ihre Dominanz.
- Reptilien nehmen in ihrer Anzahl stark ab.
- Vögel und höhere Säugetiere nehmen sehr stark in ihrer Anzahl zu.
(z.B. erste große Plazentatiere, Urinsektenfresser, pflanzenfressende Urhuftiere,
Urraubtiere usw.)
- Die gleichwarmen Säugetiere erobern auch kältere Lebensräume
- Beuteltiere werden weitgehend verdrängt.
- Doof gelaufen für den Rest der Lebewesen: Auftreten des Urmenschen! ;-)
Quartär: Zeitraum von -2 Mio. Jahren bis heute (umfasst Pleistozän und Holozän)
Pleistozän
- v.a. durch Eiszeiten Aussterben vieler Säugetierarten und vieler Pflanzenarten
- Urmensch entwickelten sich stark fort.
Holozän: (=Jetztzeit)
- Beginn vor etwa 10 000 Jahren
- heutige Tier- und Pflanzenwelt
In jedem Zeitalter liegt eine charakteristische Flora und Fauna vor.
Die Erdzeitalter in der Übersicht
Unser Planet existiert seit ca. 4,55 Milliarden Jahren. Diese Zeitspanne teilen Geologen in 4 Äonen ein. Allerdings lassen Forschungen im Gestein unseres Planeten meist nur eine Untersuchung des letzten Äons zu. Zudem war davor die Erdkruste nicht völlig erstarrt, was eine heutige Untersuchung so gut wie unmöglich macht. Die davor liegenden Äonen fasst man auch als „Präkambrium“ zusammen.
|
Äonothem |
Ärathem |
System |
≈ Beginn |
|
Phanerozoikum |
|||
|
Känozoikum |
Quartär |
2,588 |
|
|
Neogen |
23,03 |
||
|
Paläogen |
65,5 |
||
|
Mesozoikum |
Kreide |
145,5 |
|
|
Jura |
199,6 |
||
|
Trias |
251 |
||
|
Paläozoikum |
Perm |
299 |
|
|
Karbon |
359,2 |
||
|
Devon |
416 |
||
|
Silur |
443,7 |
||
|
Ordovizium |
488,3 |
||
|
Kambrium |
542 |
||
|
Proterozoikum |
|||
|
Neoproterozoikum |
Ediacarium |
635 |
|
|
Cryogenium |
850 |
||
|
Tonium |
1.000 |
||
|
Mesoproterozoikum |
Stenium |
1.200 |
|
|
Ectasium |
1.400 |
||
|
Calymmium |
1.600 |
||
|
Paläoproterozoikum |
Statherium |
1.800 |
|
|
Orosirium |
2.050 |
||
|
Rhyacium |
2.300 |
||
|
Siderium |
2.500 |
||
|
Archaikum |
Neoarchaikum |
2.800 |
|
|
Mesoarchaikum |
3.200 |
||
|
Paläoarchaikum |
3.600 |
||
|
Eoarchaikum |
4.000 |
||
|
Hadaikum |
4.600 |
||
Quelle: GNU Public Lizense, http://de.wikipedia.org/wiki/GNU_General_Public_License, Autoren siehe http://de.wikipedia.org/w/index.php?title=GNU_General_Public_License&action=history, http://de.wikipedia.org/wiki/Erdzeitalter
Zusatzinformationen:
http://de.wikipedia.org/wiki/Erdzeitalter
Brückentiere
Brückentiere sind aus der heutigen Sicht „Übergangsformen“ oder „connecting links" (manchmal auch gefundene „missing links“). Sie zeigen Merkmale zweier Gruppen, zwischen denen sie eine Zwischenstellung einnehmen (connecting links). Demzufolge hat bei ihnen im Stammbaum eine Aufspaltung in zwei Linien stattgefunden. Diese Übergangsformen zwischen zwei Tier- oder Pflanzengruppen weisen sowohl Merkmale der phylogenetisch älteren als auch jüngeren Gruppe auf.
Brückentiere, welche Merkmale zweier Gruppen aufweisen werden auch Mosaikform genannt.
Brückentiere zeigen Merkmale mehrerer Tiergruppen (=> sie hatten gemeinsame Vorfahren!)
Sie werden auch als „Missing Link“ bezeichnet, wenn es fossile Formen sind, welche eine Lücke im Stammbaum zwischen zwei Arten oder zwei Gruppen schließen.
a) Der Archaeopterix
Ein bekanntes Beispiel für ein solches Brückentier ist der Urvogel Archaeopterix. Gefunden im fränkischen Jura, lebte vor ca. 140 Mio. Jahren. Er stellt ein Bindeglied zwischen Reptil und Vogel dar.

Quelle Grafik: Public domain by Wikicommonsuser Frederik Spindler:
http://commons.wikimedia.org/wiki/Image:Archaeopteryx_Spindler2005s.jpg
Merkmale eines Kriechtiers:
Kiefer mit Zähnen, Hautschuppen, gespreizte Finger mit Krallen, Schwanzwirbelsäule
Merkmale eines Vogels:
Körperform, Federn, Flügel und Flugfähigkeit, 3 Zehen nach vorn gerichtet, der 4. Zeh ist nach hinten gerichtet (=Fersensporn).
|
„Reptilien“-Merkmale |
Vogelmerkmale |
|
|
Kopf |
kleines, einfaches Gehirn Reptilienartiger Kopf |
Vogelschädel mit großen Augenhöhlen, Schnabel vermutlich vergrößertes Kleinhirn (dient der 3D Bewegungskoordination) |
|
Rumpf |
lange, bewegliche Schwanzwirbelsäule Brustbein noch klein und ohne Kamm Wirbelform eher reptilienartig Beckenknochen sind nicht verwachsen |
Brustbein und Rabenschnabelbein |
|
Extremitäten |
3 freie, nicht verwachsene (!) Finger (ohne Endglieder) mit Krallen |
3 Finger im Flügel Die vorderen Gliedmaßen sind Flügel Oberschenkelform ist vogelähnlich |
|
Sonstiges |
nur sehr einfaches Gehirn
|
Federn und Federkleid
|
Das Archaeopterixskelett
So sah eines der vom Archaeopterix gefundene versteinerten Skelette aus:

Wo würdest Du die folgenden Tiere im Stammbaum einordnen?
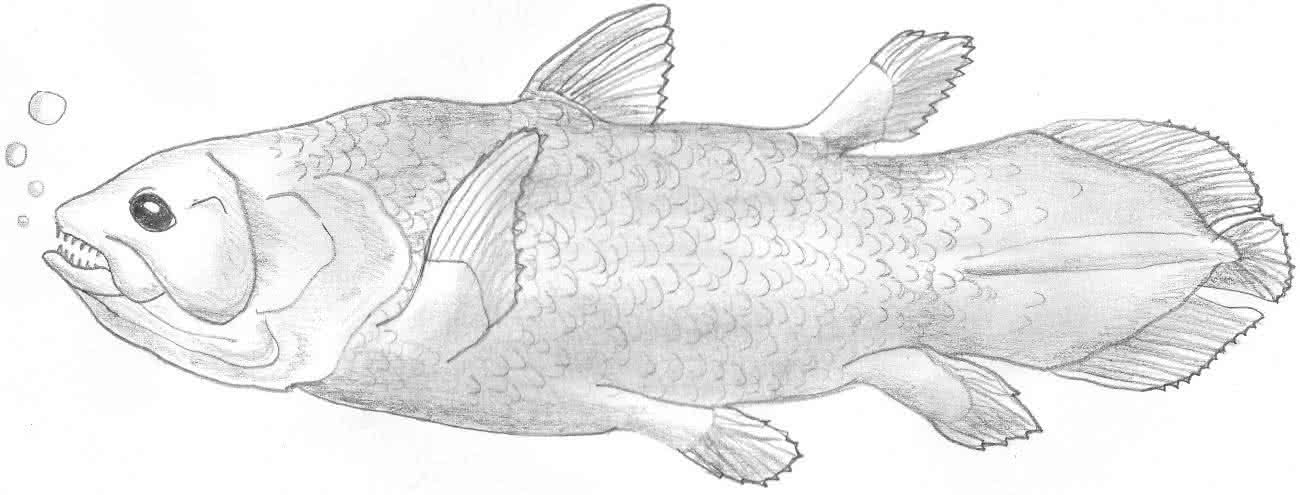
a) Quastenflosser
(ausgestorbene Art: Crossopterygier)
Lebte vor etwa 380 Millionen Jahren:

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Quelle Bild: Public Domain by Wikicommons & FishBaseartist Robbie Cada:
http://commons.wikimedia.org/wiki/Image:Coelacanth.png
{kind=link}
- lebte im Süßwasser
- sehr frühes Wirbeltier
- Hatte vier Flossen, welche schon ein wenig wie Gliedmaßen aussahen (enthielten bereits feste Knochen!).
- Mit diesen Gliedmaßen war eine plumpe und einfache Bewegung an Land möglich (z.B.: während Trockenperioden)!
- Hatte zwar Kiemen, konnte an Land aber auch über die Schwimmblase Atemgase aus der Luft aufnehmen!
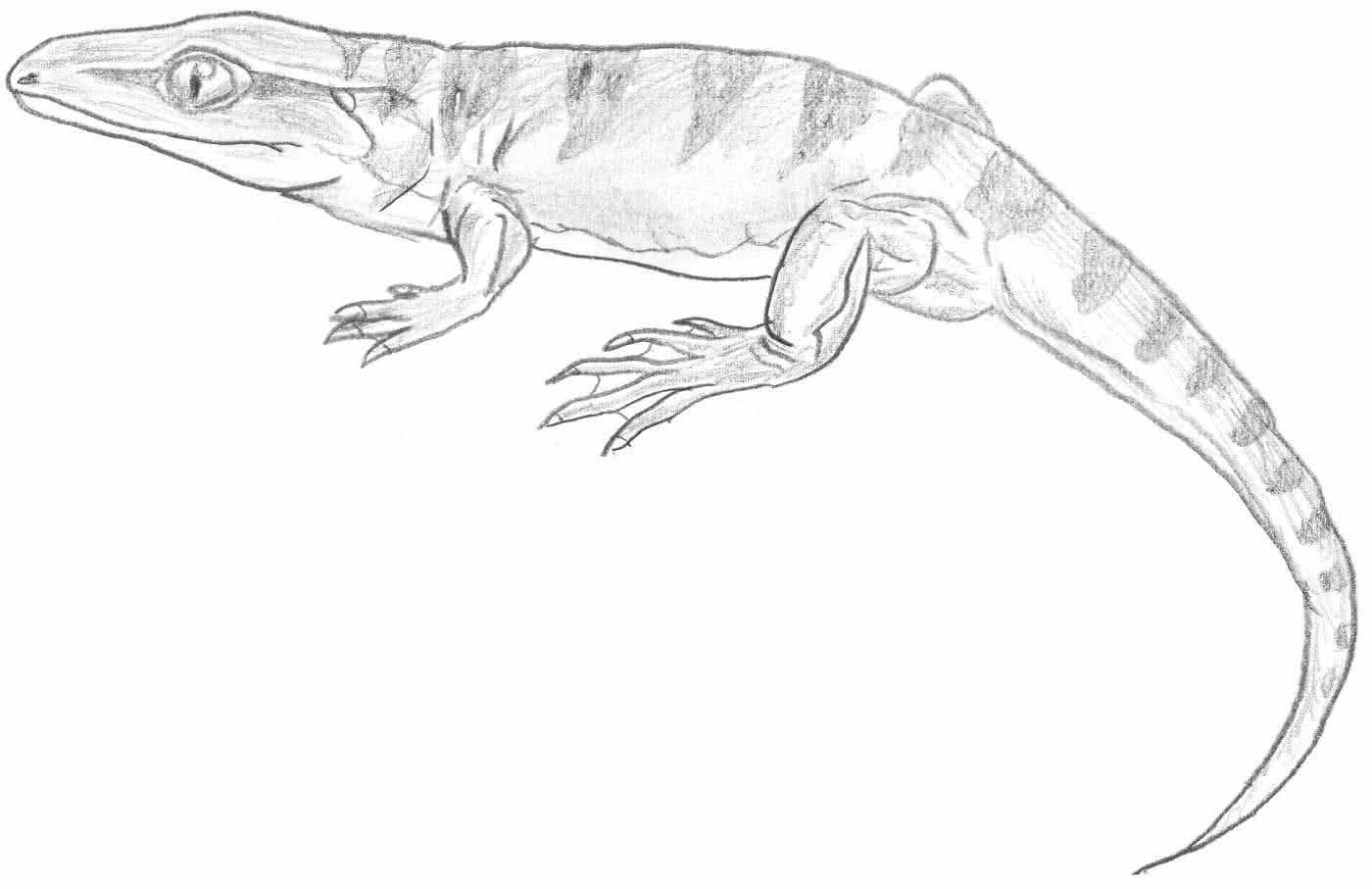
b) Ichthyostega (Devon Grönlands)
- Lebte vor ca. 350 Mio. Jahren:
- verfügte bereits über richtige Gliedmaßen mit je 5 Zehen, Schulter- und Beckengürtel, hatte allerdings noch eine Schwanzflosse (mit Schuppen bedeckt).
- „Fisch mit Beinen“: Bindeglied zwischen Fischen und Amphibien
Seitenlinienorgan - Der Körperbau ähnelte dem heutiger Schwanzlurche
- besaß noch sein Seitenlinienorgan und ein fischähnliches Gebiss.

Quelle Bild: Creative Commons Attribution 2.5 License by Wikicommonsuser ArthurWeasly: http://commons.wikimedia.org/wiki/Image:Ichthyostega_BW_2.jpg
{kind=link}
c) Seymouria (Karbon Texas)
- Lebte vor 280 Mio. Jahren:
- Bindeglied zwischen Amphibien und Reptil
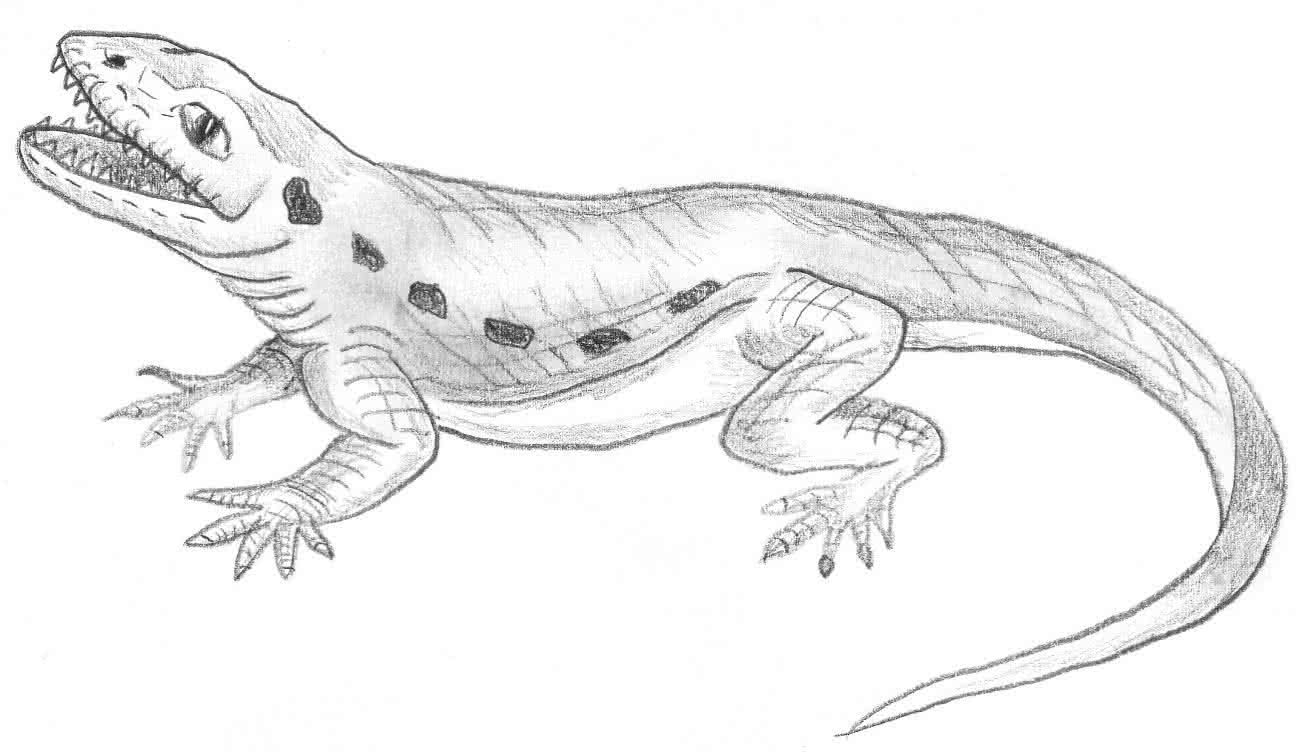
d) Cynognathus:
- Bindeglied zwischen Reptil und Säugetier (Trias)
- Reptilienmerkmale: Reptilienschwanz, Körperbau
- Säugermerkmale: Säugetierzähne, Fell
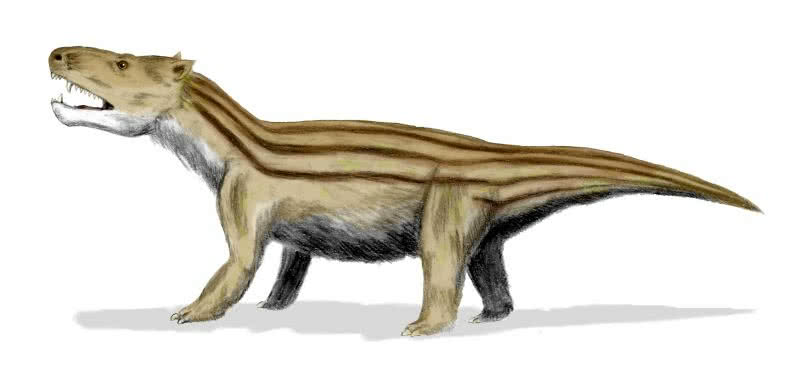
Quelle Bild: GNU Free Documentation License 1.2 & Creative Commons Attribution ShareAlike 3.0 by Wikicommonsuser Nobu Tamura - Thank you; http://en.wikipedia.org/wiki/File:Cynognathus_BW.jpg; http://en.wikipedia.org/wiki/GNU_Free_Documentation_License; http://creativecommons.org/licenses/by-sa/3.0/
{kind=link}
Merkmalszuordnung von Ichthyostega und Archaeopterix zu ihren jeweiligen Gruppen:
|
Ichthyostega |
Archaeopterix |
|
zwischen Fisch & Amphibien |
zwischen „Reptil“ & Vogel |
|
Fischgestalt |
Zähne |
|
fischähnlicher Schwanz |
Schuppen |
|
Rückenflosse |
gespreizte Finger |
|
Schwanz |
|
|
4 Beine |
Federn |
|
Knochen |
Flügel und Flugfähigkeit |
|
5 Zehen |
3 Zehen nach vorn, der 4. nach hinten (=Fersensporn) |
|
keine Kiemen |
|
|
Erstes bekannte Landwirbeltier |
Zusatzinformationen:
- http://de.wikipedia.org/wiki/Archaeopteryx
- http://de.wikipedia.org/wiki/Ichthyostega
- Foto eines Modells: http://en.wikipedia.org/wiki/Image:Archaeopteryx-model.jpg
- http://de.wikipedia.org/wiki/Quastenflosser
- http://de.wikipedia.org/wiki/Ichthyostega
- http://de.wikipedia.org/wiki/Cynognathus
- http://de.wikipedia.org/wiki/Mosaikform
{kind=link}
Lebende „Fossilien“ (= noch lebende Dauerformen)
Neopilina (Urschnecke):
- 1951 entdeckt, nur noch ein lebender Vertreter einer ehemalig größeren Gruppe!
- lebt in der Tiefsee der Meere unterhalb von 3500 m
- Vergleichbare fossile Vorformen lassen auf eine 450 Mio. Jahre lange Stammeslinie schließen.
Nautilus (Perlboot):
- Heute noch drei lebende Arten der ehemals großen Gruppe bekannt
- Merkmal: Kopffüßer mit umfangreicher Schale
- Verwandt mit Tintenfischen
- Vorkommen im südwestlichen Pazifik
- Fossile Formen kennt man schon aus dem Perm (vor 150-200 Mio. Jahren)
Quastenflosser (noch lebende Art: Latimeria chalumnae):
- 1938 erstmals rezent wiederentdeckt
- lebt heute in einer Tiefe von 150-800m im Meer (Salzwasser!) um die Komoreninseln
- einziger rezenter Vertreter der Gruppe der Crossopterygier - alle anderen sind längst ausgestorben! Deshalb dachte man auch, dass dieser seit 80 Mio. Jahre für ausgestorben sei!
Pfeilschanzkrebs (Xiphosura)
- Heute noch drei lebende Arten der ehemals großen Gruppe bekannt
- Gehört zu den Gliedertieren. Somit liegt Verwandtschaft mit Krebsen, Spinnen und Insekten vor.
- Fossile Formen kennt man schon aus dem Perm (bis vor 175 Mio. Jahren!
Schnabeltier Ornithorhynchus anatinus:
- Kloake als Auslassöffnung für Exkremente und Fortpflanzungszellen (äußere Befruchtung über Eierlegen)
- Haare am ganzen Körper
- entenartiger Schnabel
- wechselwarm
- säugt (!) seine Jungen
Zusatzinfotmation:
Der Gingko gilt als Bindeglied zwischen Nadelbaum und Laubbaum.
Übersicht über einige noch lebenden „Fossilien“
a) Pflanzen
- Ginkobaum
- Amborellaceae
- Cycads
- Horsetails Equisetum (Equisetaceae)
- Metasequoia Dawn Redwood (Cupressaceae; a borderline example, related to Sequoia and Sequoiadendron)
- Sciadopitys tree (Sciadopityaceae)
- Whisk ferns Psilotum (Psilotaceae)
- Welwitschia (Welwitschiaceae)
- Wollemia tree (Araucariaceae; a borderline example, related to Agathis and Araucaria)
b) Pilze
- Neolecta
c) Tiere
Wirbeltiere
Säugetiere
- Cypriot mouse (Mus cypriacus)
- Giant Panda (Ailuropoda melanoleuca)
- Okapi (Okapia johnstoni)
- Laotian Rock Rat (Laonastes aenigmamus)
- Monito del Monte (Dromiciops gliroides)
- monotremes (the platypus and echidna)
- Mountain Beaver (Aplodontia rufa)
- Opossums
Vögel
- Magpie-goose (Anseranas semipalmata)
- Hoatzin (Ophisthocomus hoazin)
- Coliiformess (mousebirds and relatives, 6 living species in 2 genera)
- Acanthisittidae (New Zealand "wrens")
- Broad-billed Sapayoa (Sapayoa aenigma)
- Bearded Reedling (Panurus biarmicus)
Reptilien (beide Gruppen)
- Crocodilia (crocodiles, gavials and alligators)
- Tuataras (Sphenodon punctatus and Sphenodon guntheri)
Amphibien
- Purple frog (Nasikabatrachus sahyadrensis)
Knochenfische
- Bowfin (Amia calva)
- Coelacanth (the lobed-finned Latimeria menadoensis and Latimeria chalumnae)
- Queensland lungfish (Neoceratodus fosteri)
- Sturgeons and paddlefish (Acipenseriformes)
Haie
- Frilled shark (Chlamydoselachus anguineus)
Nichtwirbeltiere
Insekten
- Mantophasmatodea (gladiators; a few living species)
- Mymarommatid wasps (10 living species in genus Palaeomymar)
- Notiothauma reedi (a scorpionfly relative)
- Orussidae (parasitic wood wasps; about 70 living species in 16 genera)
- Peloridiidae (peloridiid bugs; fewer than 30 living species in 13 genera)
- Sikhotealinia zhiltzovae (a jurodid beetle)
- Syntexis libocedrii (Anaxyelidae cedar wood wasp)
Krebse
- glypheoid lobsters (3 living species: Neoglyphea inopinata, N. neocaledonica, and Laurentaeglyphea neocaledonica)
- Triops cancriformis (a notostracid crustacean)
Schnecken
- Nautilina (e.g. Nautilus pompilius)
- Neopilina galateae, a monoplacophorid mollusc
- Ennucula superba (Nut clam)
andere Nichtwirbeltiere
- Crinoids
- Horseshoe crabs (only 4 living species of the class Xiphosura, family Limulidae: Limulus polyphemus,Tachypleus gigas, Tachypleus tridentatus and Carcinoscorpius rotundicauda)
- Lingula anatina (an inarticulate brachiopod)
- Onychophorans
- Valdiviathyris quenstedti (a craniforman brachiopod)
Zusatzinformationen:
http://de.wikipedia.org/wiki/Lebendes_Fossil
http://en.wikipedia.org/wiki/Living_fossil
Der Stammbaum der Säugetiere

Zusatzinformationen:
http://de.wikipedia.org/wiki/Säugetiere
Die drei Gruppen des Stammbaums der Säugetiere
Man kennt heute ca. 5500 Arten Säugetierarten weltweit.
Man kann sie in drei 3 Gruppen (eierlegenden Ursäuger (Protheria), die Beutelsäuger (Metatheria) und die höheren Säugetiere/ Plazentatiere (Eutheria) einteilen.
a) Ursäuger (Protheria)
Man kennt nur noch eine Gruppe innerhalb der Ursäuger. Sie umfasst fünf Arten und wird Kloakentiere genannt.
Die Kloakentiere:
Vertreter: Schnabeltier, Schnabeligel, Ameisenigel
insgesamt nur 5 Arten, alle 5 in Australien, Neuseeland und Neuguinea.
Merkmale: - legen Eier! (also nicht lebend gebärend),
- haben Milchfelder statt Milchdrüsen. - Sie lecken die Milch von Milchfeldern, da sie kein Maul oder Lippen haben (stattdessen haben sie einen Schnabel)
- An jedem Knöcheln hat das Schnabeltier einen Verteidigungssporn (Merkmal, welches nur das Schnabeltier hat)
- Kloake, welche eine gemeinsame Körperöffnung für Fortpflanzungszellen, Harnleiter und Darm ist. Man findet sie bei allen Vögeln, den meisten Reptilien und hier bei den Kloakentieren.)
_-_ameisenigel_Unterseite.jpg)
Quelle Bild: Public Domain by Brehms Tierleben,
Small Edition 1927 & Wikicommons - Thank You:
http://commons.wikimedia.org/wiki/Image:Ameisenigel_Unterseite-drawing.jpg
{kind=link}

_-_echinda_burningwell.jpg)
Bilder: Schnabeltier & Ameisenigel
Quelle Bild: Public Domain by John Lewin, State Library of NSW Quelle Bild: public domain by Wikicommons & Allan Whittome http://commons.wikimedia.org/wiki/Image:Platypus_by_Lewin.jpg http://commons.wikimedia.org/wiki/Image:Echinda_burningwell.jpg
{kind=link}
{kind=link}
Das Schnabeltier gilt als ursprüngliches Säugetier. Es zeigt noch einige Merkmale der Krichtiere (ehemals Reptilien):
- Reptilienmerkmale: eierlegend, Kloake,
- Säugermerkmale: Milchdrüsen, Haare
b) Beuteltiere/ Beutelsäuger (Metatheria)
Vertreter: Koalabär, Känguru (insgesamt ca. 320 Arten)
Merkmale:
- Der Nachwuchs verlässt sehr früh in seiner Entwicklung die Gebärmutter und wächst dann
in einem Beutel der Mutter heran.
- kleineres Gehirn als die Plazentatiere
- Löcher in der Gaumenplatte
- in der Regel 5 Schneidezähne (mit bis zu einer Gesamtzahl von 50 Zähnen!)




Alle vier Fotos fotografiert in Australien 2005 von Grit Noak - Danke
c) Plazentatiere (auch höhere Säugetiere genannt)
- Vertreter: die meisten Dir bekannten Säugetiere ;-) (spalten sich auf in Carnivora (Raubtiere) & Nagetiere & evtl. eine hypothetischen dritte Gruppe) (ca. 1200 Arten)
Aufgaben:
1. Im Stammbaum sind die drei genannten Gruppen nicht auf einer Hierarchieebene. Kann man sie dennoch vergleichen?
Zusatzinformationen:
http://de.wikipedia.org/wiki/Kloakentiere
http://de.wikipedia.org/wiki/Beuteltiere
http://de.wikipedia.org/wiki/Plazentatiere
Warum sterben Tiere aus?
a) Natürliche Auslese durch Fressfeinde und Artgenossen:
Konkurrenzausschlussprinzip: Bewohnen zwei Arten den gleichen Lebensraum und fressen sie die gleiche Nahrung (d.h. sie besetzten die gleiche ökologische Nische!), wird sich auf lange Sicht, die stärkere von beiden durchsetzten (vgl. Flugsaurier - Vögel).
b) Aussterben durch Umweltkatastrophen und schnelle Veränderungen des Biotops
Arten sterben durch Umweltkatastrophen aus (Vulkanausbruch, Eiszeit, Meteoriteneinschlag).
Die Entwicklung des Landlebens an vier Beispieltieren
Quastenflosser: frühes Wirbeltier, konnte vermutlich schon kurze Strecken über trockenes Land laufen, besaß eine einfache, nicht sehr effiziente Lunge und kräftige Flossen.
Ichthyostega: zwischen Fischen und Amphibien einzuordnen, lebte schon zeitweise an Land, hatte aber noch den langen Fischschwanz; Die Flossen waren allerdings schon in kurze, kräftige Beine mit jeweils 5 Fingern/ Zehen umgewandelt.

Gephyrostegus: lebte vorwiegend an Land, legte seine Eier aber noch im Wasser ab.
Hylonomus: lebte zwar teilweise noch im Wasser, legte aber seine Eier schon an Land ab. Diese besaßen bereit eine dünne Kalkschale.

Rekonstruktion von Tiktaalik roseae, einem Brückentier zwischen Muskelflosser-Fischen und Landwirbeltieren aus dem Oberdevon -
Quelle Bild: Public Domain by Wikicommonsuser TomCatX & Zina Deretsky, National Science Foundation: http://commons.wikimedia.org/wiki/Image:Tiktaalik_roseae_l ife_restor.jpg
Zusatzinformationen:
- http://de.wikipedia.org/wiki/Hylonomus
- Bild: http://commons.wikimedia.org/wiki/File:Hylonomus_BW.jpg
- http://de.wikipedia.org/wiki/Ichthyostega
- Bild: http://commons.wikimedia.org/wiki/File:Ichthyostega_skull.jpg
- http://de.wikipedia.org/wiki/Quastenflosser
{kind=link}
{kind=link}
Wichtig: Die heutigen Quastenflosser entsprechen nicht den Ursprünglichen! Erstens ist „Quastenflosser“ ein Überbegriff, zweitens fand eine Entwicklung seitdem statt!
Der Birkenspanner zeigt, wie Evolution funktioniert
Der Birkenspanner ist ja bereits bekannt. Er ist ein weiß-grauer Schmetterling, welcher durch seine Farbe besonders auf Birken gut getarnt ist. Er ist ein gerne verwendetes Beispiel für den Beweis der Evolution.
Eine (zufällige) Veränderung im Erbgut (= Mutation) des Birkenspanners verändert die Farbe seiner Flügel. Er ist durch diese Mutation fast braun-schwarz. Normalerweise haben solche Birkenspanner keine besonders große Lebenserwartung, da sie an den Birken leicht von ihren Fressfeinden, den Vögeln, zu erkennen sind.
Im frühindustriellen England des späten 19. Jahrhunderts färbten sich die Birken aber durch die enorme Luftverschmutzung mit Ruß sehr dunkel. Auf diesen Birken wurde die weiße Variante schneller gefressen, während die dunkle gut getarnt war. Dadurch nahm die Anzahl der dunklen Birkenspanner deutlich zu. Es liegt also ein Beispiel der natürlichen Selektion vor. Lebewesen, welches am Besten an die Natur angepasst sind, haben die besten Überlebenschancen und können somit besonders viele Nachkommen haben (Survival of the fittest).
Weitere Beispiele für das Wirken von Mutation und Selektion:
1961 wurde auf einem Bauernhof in Großbritannien eine Katze geboren, welche keine Stehohren, sondern seitlich abgeknickte Ohren hatte. Sie wurde vom Menschen weiter gezüchtet (menschliche Selektion). Es wurde eine neue Katzenrasse geschaffen: die schottische Faltohrkatze. Vermutlich im Jahre 1190 zeigte eine Rotbuche im Frühjahr ein Wachstum von roten, statt grünen Blättern. Sie wurde Blutbuche genannt. 1836 wuchs eine weitere besondere Rotbuche. Sie hatte hängende Zweige und Blätter. Sie wurde Trauerbuche genannt. Neue Buchenarten sind durch Punktmutationen aus der Rotbuche entstanden.
Aufgaben:
- Welchen Selektionsvorteil hatte der Archaeopterix gegenüber den Reptilien seiner Zeit? Warum ist er trotzdem ausgestorben?
- Das Alpenschneehuhn hat ein weißes Winterkleid. Finde einen Erklärungsweg, wie es dazu kommen konnte. Welchen Selektionsvorteil hat das Tier dadurch?
- Denke mal an die Nahrungsbeziehungen zurück: Viele Arten leben mit ihren Fressfeinden im biologischen Gleichgewicht. Welche Tiere werden aussortiert? Welchen Vorteil hat dies?
Zusatzinformationen:
http://de.wikipedia.org/wiki/Birkenspanner
Kontinentalverschiebungstheorie
Der Polarforscher und Meteorologe Alfred Wegner (1880-1930) hat eine Theorie aufgestellt, wonach sich die Kontinente nicht fest auf einer Position befinden, sondern vielmehr bewegen. Ihm fiel z.B. auf, das die westliche Küstenlinie Afrikas gut an die östliche Küste Südamerikas passen würde.
Geologische Untersuchungen zeigen auch, das beide Küsten identisches Gestein mit zum Teil in den gleichen Tier- und Pflanzenfossilien besitzen.
Als Ursache gilt, die die Erde umgebende, bewegliche Erdkruste, welche auf den darunterliegen flüssigen Magmaschichten „schwimmt“. Die Kontinente tauchen dabei ca. 30-40 km tief in die glühende Magma des Erdinneren ein.
Die Erdkruste besteht aus verschiedenen geologischen Platten, welche so gegeneinander beweglich sind. 70% der Erdkruste sind von Meeren bedeckt. Die restlichen 30% stellen heute die Landmasse der Kontinente dar.
Dabei kann man nun unterschieden zwischen den Meeresböden (aus schwerem Basalt) und den Landmassen / Kontinenten (aus leichterem Granit).
Ähnlichkeit von Merkmalen I: Übernommene Merkmale (Plesiomorphien)
Um Merkmale genau analysieren zu können, muss man ihren stammesgeschichtlichen Ursprung kennen. Dabei ist es entscheidend, Homologien von Analogien zu unterscheiden.
Plesiomorphe Merkmale sind nicht neu entstanden und schon bei vorhergehenden Arten zu finden. Diese Merkmale haben sich nicht verändert. Ein Beispiel sind die Haare des Menschen. Dieses Merkmal ist schon vorher zu finden, es taucht sogar bei allen Säugetieren auf.
=> Eine Verwandtschaft ist also nur weitläufig durch solche Merkmale zu bestimmen.
Ähnlichkeit von Merkmalen II: Homologien
Homologe Merkmale (homos gr. gleich, übereinstimmend) (bzw. seltener auch homologe Verhaltensweisen) ähneln sich in der Regel nicht.
Homologie liegt z.B. bei dem Flügel der Fledermaus und der menschlichen Hand vor. Eine weitaus schwerer zu verstehende Homologie liegt zwischen dem Wirbeltierzahn und der Haifischschuppe vor. Aber Achtung, homologe Merkmale müssen nicht gleich aussehen. Oft haben sich Aussehen und Funktion im Laufe der Evolution verändert. Ihre Gemeinsamkeit ist eher der gemeinsame Grundbauplan (wohinter dann gleiche Gene stecken).
Ein solcher Funktionswechsel kann als Anpassung an eine veränderte Lebensweise oder einen anderen Lebensraum entstanden sein. Homologe Organe können deshalb unter Umständen sehr unterschiedlich gestaltet sein! Eine ehemals gleiche Grundstruktur ist im Laufe der Zeit und der Veränderung der ökologischen Erfordernisse variiert.
Damit können homologe Organe sehr unterschiedliche Funktionen haben und somit auch sehr verschieden aussehen bzw. sehr unterschiedlich gestaltet sein!
Bekannte Beispiele sind die schon genannten Vordergliedmaßen der Wirbeltiere (z.B. als Laufbeine, als Flügel (Vögel, Flugsaurier, Fledermäuse), als Flossen (Fische, Pinguine, Ichthyosaurier, Wale), als Greifwerkzeuge (Mensch, Affen), als Grabwerkzeuge (Maulwürfe, Nacktmullen).
All diese homologen Organe haben den gleichen ursprünglichen Bauplan, welcher auf ein gemeinsames Ursprungsgen zurückgeht.
=> Homologien sind zum Beweisen von Verwandtschaftsverhältnissen der wichtigste Beleg. Liegt eine bewiesene Homologie vor, so ist die stammesgeschichtliche Verwandtschaft der verglichenen Tiere oder Pflanzen sicher bewiesen.
Merkmale, Organe oder Strukturen, welche sich auf den gleichen Grundtyp/ Grundbauplan zurückführen lassen (in Bau und Entstehung), bezeichnet man als homolog!
Homologen Strukturen liegt ein gemeinsames Erbgut zugrunde!
Liegt in Folge einer gemeinsamen genetischen Information eine Homologie vor, so ist die stammesgeschichtliche Verwandtschaft der beiden Lebewesen bewiesen!
Zur Bestimmung, ob ein Merkmal homolog ist, können Homologiekriterien dienen!
Beispiele für homologe Organe:
Die knochenähnlichen Kiemenbögen der Fische findet man bei Säugetieren noch immer. Sie befinden sich als unsere Gehörknöchelchen (Hammer, Steigbügel, Amboss) im Innenohr. Sie haben eine andere Funktion und anderes Aussehen, gehen aber auf einen gemeinsamen genetischen Ursprung zurück => homologes Merkmal.
Alle Tiere der Gruppe der Bärtierchen (=Tardigrada) verfügen über feine Stilette als kalkhaltige Mundwerkzeuge. => homologes Merkmal
Tipp: Betrachtet man nun zwei Merkmale, so ist die Wahrscheinlichkeit für eine Homologie (im Gegensatz zur vermuteten Analogie) mit zunehmender Komplexität des Merkmales größer!
Quelle Bild: Public Domain by Wikicommonsuser Alexei Kouprianov sowie Carl Gegenbaur „Grundzüge der vergleichenden Anatomie“.
2. Auflage, 1870 - Danke http://de.wikipedia.org/wiki/Bild:Gegenbaur_1870_hand_homology.png
{kind=link}
„Abgefahrenes“ Beispiel zu homologen Merkmalen:
Es gibt Gene, die man bei einer unglaublichen Vielzahl an Arten findet. Je mehr Arten über ein gemeinsames Gen verfügen, desto älter ist es stammesgeschichtlich, also desto länger ist eine Entstehung her. Ein Beispiel ist die Gruppe der so genannten Homeobox-Gene (kurz Hox-Gene). Bei Wirbeltieren hat diese Gengruppe unter anderem die Aufgabe die Augen auszubilden (aber auch andere Körperabschnitte). Man findet diese Gengruppe sogar bei Drosophila melanogaster (=Fruchtfliegen), welche ja ganz andere Augen haben: Komplexaugen anstelle von Linsenaugen! Es kann als generelle Regel gelten, dass im Laufe der Evolution nicht zweimal das gleiche Gen neu erfunden wird (nur sehr wenige Ausnahmen!).
Bei dem blinden Schwanzlurch „Grottenolm“ ist diese Gengruppe auch vorhanden, obwohl er nicht über Augen verfügt. Vermutlich hat es eine andere Aufgabe. Vermutlich ist es energetisch effizienter, nicht alles immer neu zu erfinden, sondern Vorhandenes abzuwandeln.
Weitere Homologiebeispiele
- Dem Spritzloch / Atemloch der Haie liegt die gleiche genetische Herkunft wie unserem Mittelohr zugrunde.
- Die Kiemenbögen der Fische sind homolog zu den einigen Teilen des Kiefergelenkes der Reptilien und zu den Gehörknöchelchen Hammer und Amboss der Wirbeltiere.
- Ähnliches gilt für die Schwimmblasen der Fische, welche vom Ursprung her als Vorläufer der Lungen von Amphibien, Reptilien und Säugetieren gesehen werden können.
- Die Federn der Vögel und die Reptilienschuppen sind homolog
Homologe Organe bei Pflanzen
- Auch bei Pflanzen können wir homologisieren. Betrachtet man Keimblätter, Laub-, Blüten- und Staubblätter einer Pflanze, so erkennt man deren Homologie. Man spricht von Blattmetamorphosen, zu denen auch Blattdornen und Blattranken gehören.
- Blüte und Blütenstand: Bei Linum flavum (hat eine Blüte ) und Lindheimera texana (eine Asteracea, Korbblüter mit Blütenstand). Hier sieht man das die Scheinblüte zwar analog ist, aber die Einzelblüten des Blütenstands tatsächlich zu den Blüten homolog sind. Bei dem hier gewählten Artenpaar sind die Blüte bzw. der Blütenstand etwa gleich groß.
Ähnlichkeit von Merkmalen III: Analoge Merkmale
Betrachtet man nun ein Merkmal bei verwandten Arten, so muss es nicht zwangsweise auf identisches Erbgut zurückgehen. Ein Beispiel ist der stromlinienförmige Körper mit Flossen der Fische und der Seehunde bzw. der Körperbau des Wales. Es liegen unabhängige Entwicklungen vor, da die Vorfahren der Seehunde und der Wale an Land lebten und somit eine andere Gestalt hatten. Man nennt solche Merkmale, welche sich sehr ähnlich sind, aber auf unterschiedliche Entwicklungen und Gene zurückzuführen sind, analoge Merkmale.
Ursache ist in der Regel eine konvergente Evolution durch die ähnliche (also analoge) Organe entstanden sind.
Beispiele für analoge Organe und Strukturen:
- Saumflossen bei Kalmaren und Fischen
- vierkammrige Herz bei Säugetieren und Vögeln
- Insektenbein und Wirbeltierbein
- Insektenflügel und Vogelflügel
- Linsenaugen von Tintenfischen und Wirbeltieren
Analogien (ana gr. gemäß, logos gr. Wort, Vernunft) sind so etwas wie „Fallen“ für Phylogenetiker. Wenn man zwei Arten vergleicht und beide haben eine Struktur, welche auf den ersten Blick gleich ist (was dann bedeuten würde, dass Verwandtschaft vorliegt), so kann es sein, dass in Wirklichkeit die Strukturen beider Tiere zwar gleich aussehen und die gleiche Funktion erfüllen, aber einen verschiedenen Bauplan haben und verschiedener Herkunft und somit genetisch unterschiedlich sind. Oft merkt man erst bei genauer mikroskopischer Betrachtung, dass zwei vermeintlich gleiche Organe einen unterschiedlichen Grundaufbau haben.
Organe oder Strukturen, die die gleiche Funktion erfüllen, aber einen im Grunde verschiedenen Bauplan haben (und somit verschiedene Gene zugrunde liegen) und verschiedener Herkunft sind, bezeichnet man als analog.
Problematik: Homologe Organe können trotz eines gemeinsamen Ursprungs sehr verschieden gestaltet sein. Analoge Organe hingegen sind aufgrund ihrer gleichen Funktion oft sehr ähnlich! Dies erschwert eine Unterscheidung enorm. Der Grad der Übereinstimmung ist manchmal so groß, dass analoge Organe fälschlicherweise für homolog (bzw. plesiomorph) gehalten werden und erst genetische Untersuchungen eindeutige Klarheit bringen können.
Beispiel für analoge Merkmale: Das Linsenauge des Menschen und das Linsenauge des Tintenfisches sehen sich sehr ähnlich (Augenlider, Iris, Linse, Netzhaut usw.). Beide Linsenaugen haben die gleiche Funktion und einen fast identischen Aufbau (das Wirbeltierlinsenauge ist allerdings invers in Bezug auf seine Netzhaut, das der Tintenfische evers!).
Erst eine Untersuchung der Ontogenese eines Tintenfisches zeigt, dass dessen Linsenauge aus einem anderen Keimblatt entstanden ist. Eine solche Untersuchung zeigt dann, dass die Netzhaut und Pigmentschicht des Wirbeltierauges bei der Entwicklung aus einer Vorstülpung des Zwischenhirns entstehen. Beim Tintenfisch hingegen wird die Netzhaut aus Oberhaut gebildet. Sie ist also ein Teil der tierischen Epidermis!
Daraus leiten sich konstruktionsbedingt morphologische Unterschiede ab:
- Die Wirbeltiernetzhaut ist mehrschichtig.
- die Wirbeltiernetzhaut ist invers, d.h. ihre Sehzellen sind vom Lichteinfall weggewandt
- die Tintenfischnetzhaut ist hingegen einschichtig und dem Licht zugewandt, als evers.
Analoge Organe sind äußerlich sehr ähnliche Merkmale, welche die gleiche Funktion haben, aber unabhängig voneinander entstanden und nicht auf gemeinsame Gene zurückzuführen sind. Ursache ist eine konvergente Entwicklung.
Konvergente Entwicklung = Anpassung nicht-homologer Organe an die gleiche Funktion (aufgrund sehr ähnlicher Umweltbedingungen / ökologischer Nischen).
Konvergenz als Ursache für analoge Merkmale
Ursprünglich verschieden gestaltete Organe oder Strukturen zweier nicht eng verwandter Lebewesen werden im Laufe der Evolution aufgrund ihrer Anpassung an die gleiche Umwelt einander zunehmend ähnlicher. Solche analogen Merkmale entstehen also als Anpassung nicht-homologer Organe an ähnliche Umweltbedingungen bzw. ähnliche Erfordernisse. Diese Ausprägung gleicher Funktionen bezeichnet man als konvergente Entwicklung.
Besonders eindrucksvoll sind Konvergenzen, wenn ganze Organsysteme oder der Gesamthabitus der Tiere/ Pflanzen davon betroffen sind.
Beispiele:
- Fischgestalt oder Wurmgestalt als „Lebensformtyp“
- Stammsukkulenz bei verschiedenen Pflanzen
- Ranken:
Blattranken (Erbse)
Sprossranke (Weinstock, Passionsblume)
Wurzelranke (Vanille) - Dornen:
„Blattdornen“ (Rose, Kaktus, Berberitze) => werden Stachel genannt
Sprossdornen (Schlehe, Weißdorn)
Wurzeldornen (Palmen)
Homologien erklären die Biogenetische Grundregel (Haeckel):
Ernst Haeckel beobachtete Homologien von Embryonalstadien. Er machte folgende Beobachtungen:
1. Die Embryonalstadien der Wirbeltiere zeigen vor allem in den sehr jungen Stadien auffallende Übereinstimmung in der Gestalt.
2. Einige Merkmale findet man bei allen Wirbeltieren als Anlage. Dazu gehören u.a. die Kiemenbögen.
3. Einige Tiere zeigen besondere Anlagen, welche eigentlich Entwicklungsrudimente darstellen. So fand Haeckel bei den sonst zahnlosen Bartenwalen embryonale Zahnanlagen. Diese werden im Laufe der Entwicklung allerdings zurückgebildet und verschwinden dann ganz.
Auch die sonst beinlose Blindschleiche, welche ja zu den Eidechsen gehört, bildet in der Embryonalentwicklung Anlagen von Vordergliedmaßen. Bei der adulten Blindschleiche ist davon aber nur noch der Beckengürtel erhalten. Die Vordergliedmaßen haben sich dann wieder zurückgebildet.
=> Ernst Haeckel 1866: Biogenetisches Grundgesetz:
Die embryonale Entwicklung eines Lebewesens (=Ontogenese) ist die unvollständige und schnelle Wiederholung der Stammesentwicklung (=Phylogenese).
Sie zeigt einen beschleunigten Verlauf der Stammesentwicklung.
Ein Tier „klettert“ also gewissermaßen an seinem eigenen Stammbaum empor, wobei die Embryonalstadien die aktuellen Zustandsbilder der einzelnen Stämme zeigen
Belege beim Menschenembryo:
- Kiemen
- eine fischähnliche 1. embryonale Stufe
- Kleinkinder sind Außensohlengänger.
- embryonales Haarkleid
- kleine Schwanzanlage
Kritik an Haeckel:
- Ein Teil der auftretenden Bildungen stellen Eigenanpassungen des Keimes dar
(z.B. Eihäute, Dottersack). - Es werden nicht alle (!) Stadien der Phylogenese durchlaufen, auch sind nur einzelne Organe betroffen.
- Es werden keine Adultstadien rekapituliert, sondern nur Organanlagen.
=> Umformung zur Biogenetischen Grundregel:
In der Embryonalentwicklung eines Lebewesens werden auch Merkmale seiner stammesgeschichtlichen Vorfahren angelegt, die dem Erwachsenenstadium fehlen.
Erklärung
Erhaltung älterer Bauplanmerkmale
Zugrundeliegende Gene können nicht einfach verschwinden.
=> Bedeutung von Larvenstadien: Entdeckung von Verwandtschaftsbeziehungen, die durch den Vergleich der adulten Tiere nicht möglich wäre.
Belege für die biogenetische Grundregel nach Haeckel

Haeckels Zeichnungen von 1892 als Beleg für seine ontogenetische Grundregel. Heute weiß man, dass die Embryonen nicht ganz exakt sind. Es scheint, als hätte der Meister die Ähnlichkeiten überbetont.
Quelle Bild: Public domain by Wikicommons sowie Romanes's 1892 copy of Ernst Haeckel's allegedly fraudulent embryo drawings (this version of the figure is often attributed incorrectly to Haeckel). http://commons.wikimedia.org/wiki/Image:Haeckel_drawings.jpg
{kind=link}
Merke:
Ontogenese = Keimesentwicklung
Phylogenese = Stammesentwicklung
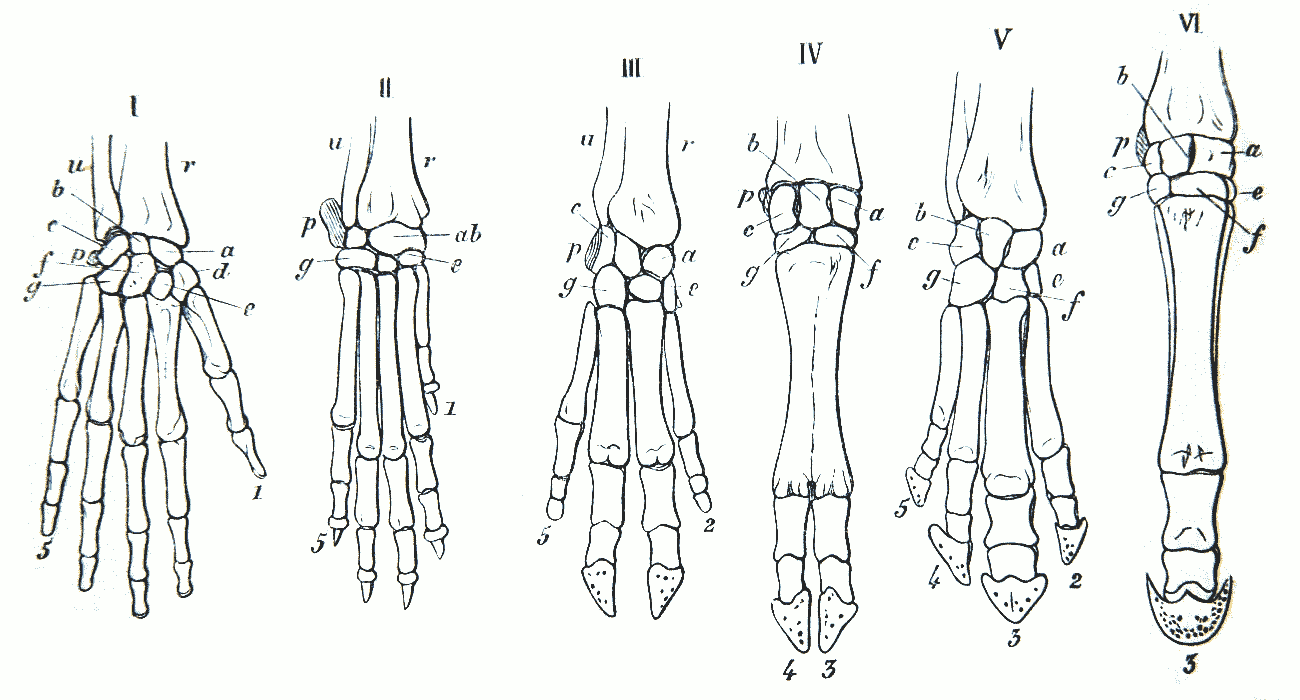
Homologievergleich des Armskeletts

120 Salamander
121 Schildkröte
122 Krokodil
123 Vogel
124 Fledermaus
125 Wal
126 Maulwurf
127 Mensch
Quelle: Public Domain by Wilhelm Leche, 1909 - thank you; http://commons.wikimedia.org/wiki/Image:Arm_skeleton_comparative_NF_0102.5-2.png
{kind=link}
Vergleich der homologen Knochen mit analogen Fluganpassungen
Man sieht die Vordergliedmaßen von drei Wirbeltiergruppen mit analog gebildeten Flügeln:
1) Flugsaurier (Pterosauria):
Die Flügel werden vom 4. Finger getragen.
2) Fledertier (Chiroptera):
Die Flügel werden vom 2. bis 5. Finger getragen.
3) Vogel (Aves):
Die Flügel werden wesentlich vom 2. Finger getragen.
Quelle Bild: Public domain by Wikicommonsuser Lauranrg & John Romanes': Darwin and after Darwin - http://commons.wikimedia.org/wiki/Image:Homology.jpg
Homologieforschung durch Homologiekriterien
Der Nachweis von Homologien zum Erstellen von Verwandtschaften ist schwierig. Um sie eindeutig nachweisen zu können, benötigt man verschiedene Kriterien.
a) Kriterium der Lage
Strukturen sind dann homolog, wenn sie trotz ihrer unterschiedlichen Ausprägung in einem vergleichbaren Gefügesystem die gleiche Lage einnehmen.
Beispiele:
Bei Insekten sind die verschieden gestalteten Insektenbeine homolog, da sie alle am Thorax (also der Brust) des Tieres ansetzen. => Kriterium der Lage erfüllt.
Die Schwimmblase der Fische und die Lungen der Landwirbeltiere sind homolog, da sie aus dem gleichen Keimblatt entstehen und die gleiche Position im Körper einnehmen.
=> homolog trotz unterschiedlicher Form!
Die Homologie von Vogelflügel und Maulwurfsbein wird also durch eine vergleichbare Lage im Skelett beider Tiere begründet.
b) Kriterium der Kontinuität (Stetigkeit)
Trifft das Kriterium der Lage nicht zu, so kann dennoch Homologie vorliegen, wenn sich trotz unterschiedlicher Lage die beiden Strukturen durch Zwischenformen (auch Fossilen), die je untereinander Schritt für Schritt homologisierbar sind, verbinden lassen.
Beispiele:
- Die Halsschlagader von Säugetieren kann aufgrund von Embryonalstadien (z.B. Kiemenspalte beim Menschen) mit den Kiemenbogenarterien der Fische als Homologie verbunden werden.
- Die Schwimmblase der Fische und die Lungen der Landwirbeltiere sind homolog, da sich die Entwicklung Schritt für Schritt von den Amphibien über die Sauropsiden nachvollziehen lässt.
- Die Gehörknöchelchen der Landwirbeltiere sind über Zwischenstufen aus ersten Kiemenbögen der Fische entstanden.
- Die Griffelbeine der Pferde sind mit den Mittelhandknochen anderer Wirbeltiere homolog, weil man durch fossile Funde zeigen kann, dass Urpferde eine schrittweise Reduktion der Zehen durchlaufen haben. (Griffelbeine sind rudimentären Mittelhand- bzw. Mittelfußknochen des 2. und 4. Fingers. Man findet sie nur beim Pferd. Sie liegen hinter dem Hauptknochen, dem Röhrbein, welches dem aus Mittelhand- bzw. Mittelfußknochen des 3. Fingerstrahls gebildet wird.)
c) Kriterium der spezifischen Qualität
Treffen die beiden vorherigen Merkmale nicht zu, so kann dennoch Homologie vorliegen, sofern zwei ähnliche Strukturen in mehreren Sondermerkmalen übereinstimmen. Entscheidend für das Homologiekriterium der spezifischen Qualität ist also der „innere Aufbau“ eines Merkmals. Stimmt er bei beiden Arten in zahlreichen Einzelheiten überein, kann Homologie vorliegen.
Beispiel:
Die Haifischschuppen (im Englischen interessanterweise „denticals“ genannt) und der Wirbeltierzahn haben den gleichen inneren Aufbau.
Beim Hai nennt man diese Schuppen auch Placoidschuppen. Sie treten bei allen Knorpelfischen auf und bedecken deren Körper. Man kann sie deutlich fühlen, wenn man mit der Hand über die Haut eines Haies streift. Sie fühlt sich scharf, in etwa wie Sandpapier, an.
Weitere abgewandelte Placoidschuppen sind zu finden bei:
- dem Dorn bei Dornhaien
- der Säge der Sägefische
- dem Schwanzdorn beim Stachelrochen
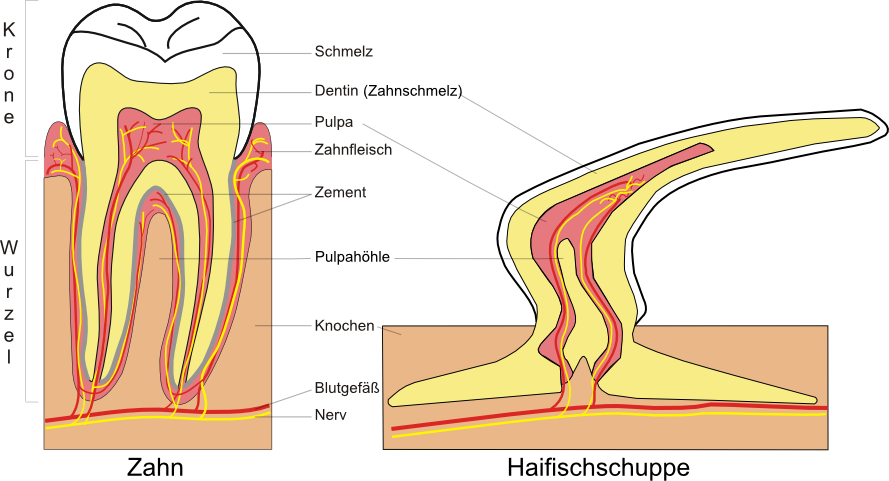
Quelle Zeichnung Zahn: Public Domain von Wikicommonsuser Uwemuell - http://commons.wikimedia.org/wiki/Image:Schematic_section_tooth.svg - Danke
Zusatzinformationen:
http://de.wikipedia.org/wiki/Homologie_%28Biologie%29#Homologiekriterien
http://de.wikipedia.org/wiki/Placoidschuppe
http://de.wikipedia.org/wiki/Ganoidschuppe
http://de.wikipedia.org/wiki/Kiemenbogen
http://en.wikipedia.org/wiki/Pharyngeal_arch
http://de.wikipedia.org/wiki/Gehörknöchelchen
http://en.wikipedia.org/wiki/Ossicles
Homologie des Verhaltens
Auch Verhalten wird vererbt. Dies gilt insbesondere für Instinkthandlungen. Aber auch hier kann Verhalten zwischen zwei Arten gleich sein, analog oder homolog!
Homologe Verhaltensweisen liegen dann vor, wenn sie sich auf dieselbe Grundhandlung zurückführen lassen. Je mehr Übereinstimmungen, desto enger die Verwandtschaft zwischen den Arten.
z.B.:
- Das ritualisierte Scheinputzen bei der Einleitung der Balz verwandter Entenarten ist sehr ähnlich (Stockerpel, Knäckerpel, Mandarinerpel).
- Futterlocken / Balzbewegung bei Hühnern und dem Fasan
- Greifreflex bei Menschen- und Affenbabies
- Hochziehen der Schultern bei Lärm
Die Warnrufe verschiedener Vogelarten (Eichelhäher, Amsel, Krähe usw.) sind ein Beispiel für analoges Verhalten.
Widersprüche zwischen homologem Aussehen und homologem Verhalten?
Mauersegler (leben auch in der Stadt) werden oft mit Schwalben (gehören zu den Sperlingsvögeln, leben nicht in größeren Städten) verwechselt. Tatsächlich sind sie nicht direkt miteinander verwandt.
Zusatzinformationen:
http://de.wikipedia.org/wiki/Mauersegler
http://de.wikipedia.org/wiki/Schwalben
http://de.wikipedia.org/wiki/Konvergenz_%28Biologie%29
Schwalben: Es gibt 75 Schwalbenarten:



Hinweise auf die Entwicklung der Arten: Rudimentäre Organe
Was haben der Blinddarm und das Steißbein gemeinsam? Ganz einfach, beides sind im Grunde überflüssige Organe, welche bei unseren Vorfahren eine Funktion hatten, aber dem Menschen nicht dienlich sind. Solche Organe, welche sich im Laufe der Evolution zurückgebildet haben, aber noch teilweise vorhanden sind, nennt man rudimentäre Organe.
Rudimentäre Organe sind Organe, Spuren oder Überbleibsel in rezenten Lebewesen, die, verursacht durch eine Änderung der Lebensweise einer Art, im Laufe der Stammesentwicklung funktionslos wurden, unnötig sind und sich zurückbildetet haben.
Die Rückbildung geschieht nicht durch Nichtgebrauch, wie z.B. bei Muskeln, sondern durch fehlende Selektion.
Beispiele beim Menschen:
- Schwanzwirbelsäule bzw. Steißbein (Rest eines früheren Schwanzes)
- Wurmfortsatz (enthielt bei unseren Vorfahren zelluloseverdauende Enzyme, heute Teil des Lymphsystems, unterstützt somit in geringem Maße das Immunsystem).
- ausgeprägter Eckzahn und verkümmerte Weisheitszähne (Eckzähne dienten der Einschüchterung von Rivalen und dem Verzehr von rohem Fleisch.)
- Rest der Augennickhaut (diente als drittes Augenlid bei verwandten Säugetieren)
- Funktionslose Muskeln der oberen Ohrmuscheln (dienten zur Bewegung und Ausrichtung der Ohren - bei vielen Affenarten noch zu finden)
- Körperbehaarung beim Menschen (diente als Wärmeisolierung)
- zurückgebildete Schwimmhäute
- Darwin'scher Ohrhöcker (Spitze der Säugerohren)
- Brustwarzen der Männer (vermutlich!)
- Verhaltensrudiment: Drohgebärden bei Mensch, Menschenaffe und Hirsch
Beispiele bei anderen Tieren
- Hinterextremitäten und Beckengürtel der Bartenwale (Blauwal)
- Wale sind ehemalige Landsäugetiere, welche sekundär ins Wasser zurückgekehrt sind.
- Reste des Ober- und Unterschenkels sind beim Grönlandwal zusätzlich vorhanden.
Weitere Anpassungen des Walkörpers an das Wasserleben:
- Stromlinienform
- Das Ende der Wirbelsäule wird zur Schwanzflosse umgewandelt. Sie dient so dem Hauptantrieb des Wals.
- Umbildung der Vorderextremitäten zu Schwimmflossen
- Zurückbildung des Felles, stattdessen Ausbildung einer Fettschicht
- Ersatz der Zähne durch Barten (Gewicht)
- Reduzierung der Halswirbel
- Die Blindschleiche zeigt einen vollständigen Schultergürtel sowie Reste eines Beckengürtels
- Beim Pferd sind die Griffelbeine als rudimentäre Mittelfußknochen vorhanden.
- Rückbildung der Flügel beim Pinguin
- Reste des Ober- und Unterschenkels beim Grönlandwal
- Reste von Hinterextremitäten bei Riesenschlangen
- Abstufungen der Augenrückbildung bei Höhlentieren (z.B. Blindmulle, Fledermäuse usw.)
- Gehäusereste bei Nacktschnecken
- Augen beim Maulwurf
- Riesenschlangen haben noch Reste der Hinterextremität und des Beckens.
- Kiwi und Strauß haben noch kleine stummelförmige Flügel, obwohl sie flugunfähig sind.
- Schildkröten haben noch Rippen, welche mit dem Panzer verwachsen sind. Obwohl diese Rippen somit unbeweglich sind und keine Rippenatmung (Brustkorbatmung) möglich ist, haben einigen Arten seit 200 Mio. Jahren immer noch Zwischenrippenmuskeln.
- Bartenwale haben noch Reste von Zähnen im Kiefer, obwohl sie eigentlich Krill filtrieren.
- Seelöwe und Walross haben noch Nägel an den Flossen.
- Beuteltierembryonen bilden noch einen Eizahn aus (mit dem sich einfachere Tiere aus dem Kalkei befreiten).
- Viele flugunfähige Laufkäfer haben Reste von Hautflügeln unter ihren verwachsenen Flügeldecken.
Hinweise auf die Entwicklung der Arten: Atavismen
Sind Organe bei einer Art vollständig zurückgebildet, treten aber vereinzelt noch bei einigen Mitgliedern dieser Art auf, so spricht man von einem Atavismus.
Die Ausbildung eines ursprünglichen Organs, welche als Rückschritt in der Stammesgeschichte zu erklären ist, nennt man Atavismus.
Beispiele sind beim Menschen die Ausbildung eines Schwanzes, mehrerer Brustwarzen oder auch einer starken Ganzkörperbehaarung (fellähnlich).
Der Grund, warum Atavismen als Rückschritte zu früheren Evolutionsstufen führen, liegt in der falschen Aktivierung von Genen. Viele unserer Gene sind phänotypisch inaktiv (und rezessiv). Meist werden sie vom Organismus zusätzlich blockiert. In sehr seltenen Fällen werden sie aber dennoch während der Ontogenese aktiviert, was dann zur Ausbildung von Atavismen führt.
Beispiele:
- Sehr selten werden dreizehige Pferde geboren.
- Fruchtfliegen (Drosophila) entwickeln selten häutige Hinterflügel.
- Das Wegerichgewächs „Löwenmäulchen“ hat Lippenblüten, selten treten radiärsymmetrische Blütenformen auf.
- überzählige Brustwarzen längs der Milchleiste bei Menschen
- Schwanzanlage als Steißbeinverlängerung beim Menschen

Ein bekanntes Beispiel dafür, welches wissenschaftlich allerdings noch nicht 100%ig abgesichert ist, sind das neandertalerähnliche Aussehen und Verhaltensweisen von Türstehern. *g*
Ursachen:
=> Störungen in der Embryonalentwicklung
=> Rückmutation (ungewollte Rückkreuzung)
Aufgaben:
1. Was ist der Unterschied zwischen einem rudimentären Organ und einem Atavismus?
2. Wie würdest Du die in seltenen Fällen bei menschlichen Neugeborenen auftretenden „Kiemen“ interpretieren?
Zusatzinformationen:
http://de.wikipedia.org/wiki/Atavismus
http://de.wikipedia.org/wiki/Rudiment
http://en.wikipedia.org/wiki/Vestigiality
Hinweise auf die Entwicklung der Arten: Zeugnisse aus der Paläontologie
Verwandtschaft kann durch Vergleich heute lebender Organismen mit fossilen Formen geschehen (Paläontologie). Die Kombination beider liefert Beweise für die Entwicklung der Arten. So zeigen Zwischenformen oft einen gemeinsamen Ursprung von heute lebenden (rezenten) Arten.
Fossilien = chemisch oder physikalisch (z.B. versteinerte) konservierte, vorzeitliche Lebewesen oder Reste und Spuren davon!
Voraussetzung für Fossilisation:
Rasche Sedimentation
- Knochenfunde (Knochen und Zähne)
- Sedimentabdrücke (Fußabdrücke usw)
- echte Versteinerungen
Lebewesen muss über Hartteile verfügen oder konserviert sein in
- Dauerfrostböden => Eismumien (z.B. Mammutfunde)
- fossilen Baumharzen (Bernstein)
- Einbettung in Ölschiefer (Burgeus-Schiefer)
- Mooren durch Gerbsäuren (Moorleichen)
Nachweis von Verwandtschaft: Die Präzipitin-Reaktion
Die Präzipitin-Reaktion erlaubt Rückschlüsse auf den Verwandtschaftsgrad zwischen zwei Arten. Dazu wird die Proteinähnlichkeit auf serologischem Weg durch Antigen-Antikörper-Reaktion untersucht. So sind Aussagen über den Verwandtschaftsgrad zwischen beispielsweise Mensch und Schimpanse möglich.
Einem Testtier (z.B. einem Kaninchen) wird Blutserum des Menschen gespritzt. Das Testtier sollte nicht näher mit dem zu untersuchenden Tier verwandt sein! Im Testtier läuft nun eine Immunabwehrreaktion gegen die ihm körperfremden Antikörper (also das Humaneiweiß). Nach ca. 6-8 Wochen wird dem Testkanninchen nun Blut entnommen und daraus das Serum gewonnen (= Antihumanserum!).
Dieses Antihumanserum enthält nun Antikörper gegen Humaneinweiße. Kommt das Serum nun mit Humaneiweißen in Kontakt, so flockt es aus und wird trüb (aufgrund der Verklumpungsreaktion der Antikörper).
=> Ausfällung des menschlichen Eiweißes (= Agglutination) zu 100%
Gibt man nun Serum in das Blut eines anderen Tieres, so kommt es entsprechend der Verwandtschaft ebenfalls zu einem Verklumpen, entsprechend des Verwandtschaftsgrades.
=> abgeschwächte Ausfällung
Denn je höher die Verwandtschaft, desto eher befinden sich im Tier auch gemeinsame Eiweiße!
Der Grad der Ausfällung entspricht also dem Grad der Verwandtschaft.
- Mensch (100%) <–> Schimpanse (85%)
- Mensch <–> Gorilla (64%)
- Mensch <–> Orang Utan (42%)
Die Präzipitinreaktion ist eine Eiweißfällreaktion, welche grob Rückschlüsse auf den Verwandtschaftsgrad zwischen zwei Arten erlaubt.
oben: Kaninchenserum mit Antikörpern
=> Ausfällung (Niederschlag = Präzipitin)
Präzipitintest: Anti-Humantestserum
- Menschen 100 % Ausfällung
- Schimpansen 85 %
- Gorilla 64 %
- Orang Utan 42 %
- Pavian 29 %
- Rind 10 %
- Rotwild 7 %
- Pferd 2 %
Nachweis von Verwandtschaft: die Molekularbiologie zeigt die Homologie von Makromolekülen
Erinnere Dich: Basensequenz der DNA => AS-Sequenz => Protein
1. Möglichkeit: Basensequenzanalyse:
Wird durch eine Gelektrophorese von zwei Tieren die DNA sequenziert und dann die Sequenz verglichen, so kann man sagen, dass aufgrund der Universalität des genetischen Codes eine hohe Übereinstimmung des Codes als Beweis für einen hohen Verwandtschaftsgrad gilt.
2. Möglichkeit: DNA-Hybridisierung:
Der DNA-Doppelstrang von zwei zu vergleichenden Lebewesen wird getrennt und dann werden beide Einzelstränge zusammengefügt. So kann man schnell erkennen, welche Abschnitte komplementär sind.
Je höher die Anzahl komplenetärer Basesequenzen,
desto schneller geschieht die Bildung des neuen DNA-Hybid-Doppelstranges.
desto hitzebeständiger ist der neue Doppelstrang => Eine höhere Schmelztemperatur ist ein Indiz für hohen Verwandtschaftsgrad.
Methoden der Altersbestimmung
Grob gesagt gibt es zwei Methoden der Altersbestimmung: die relativen und die absoluten Methoden.
a) Relative Altersbestimmung (Stratigraphische Methode):
Man versucht, das Alter eines Fossils dadurch zu bestimmen, indem man das Alter der Fundstelle (bzw. der geologischen Schicht) bestimmt (was durch spezielle andere Funde bzw. dokumentierte Schichtungen leichter ist). Hintergrund ist, dass das Fossil entsprechend seines Alters in eine bestimmte, zu der Zeit sich bildende Gesteinsschicht eingebettet ist.
Verfälschend wirken hier vor allem das Wetter, Erdbeben sowie die Plattentektonik, welche zu einem falschen Fundort führen können.
Kommen Fossilien in einer bestimmten Schicht besonders häufig vor, werden sie auch von Archaeologen als „Leitfossilien" bezeichnet:
- Trilobiten für das Paläozoikum
- Ammoniten für das Mesozoikum
Findet man also Fossilien mit Leitfossilien zusammen, so kann deren Alter leicht bestimmt werden, auch wenn man sonst nicht so viel über das Gestein weiß.
Fluortest
In fluoridhaltigen (F-) Böden und auch in fluoridhaltigen Wasser werden Hydroxidionen gegen Fluorid ausgetauscht. Je länger ein Tier in einem solchen Lebensraum lebt, desto höher ist die aufgenommene Menge. So kann man indirekt das relative Alter in einem gemeinsamen Fundgebiet berechnen.
b) Absolute Altersbestimmung:
1. Die 14C-Methode = Radiokarbonmethode:
Diese Methode wurde 1948 von Willard Frank Libby entdeckt. Er erhielt dafür 1960 den Nobelpreis.
Bei dieser Methode wird das Alter eines Fossils dadurch bestimmt, dass der Anteil des noch enthaltenen 14C-Isotops gemessen wird.
Dazu muss man verstehen, dass es in der Atmosphäre neben dem Kohlenstoff mit der Massenzahl 12 (12C) auch ein instabiles 14C-Isotop gibt. Es entsteht aus dem normalen Stickstoff durch kosmische Strahlung in 30km Höhe in der Atmosphäre. Sein Anteil in der Atmosphäre ist konstant! Dieses 14C-Isotop wird durch die Photosynthese in alle Pflanzen eingebaut und gelangt so in jedes Tier.
=> Jedes Lebewesen nimmt das 14C-Isotop mit der Nahrung auf, solange es lebt!
Das 14C-Isotop zerfällt unter Aussendung von Betastrahlung allerdings in Stickstoff (=14N). Das Verhältnis von 12C zu 14C ist seit Millionen von Jahren gleich geblieben!
Stirbt ein Lebewesen, kann es kein 14C mehr aufnehmen. Der vorhandene Anteil im Körper nimmt nun durch den Betazerfall stetig ab. Die Halbwertszeit liegt bei ca. 5.730 Jahren, also ist nach ca. 6000 Jahren die Hälfte des 14C-Isotops verschwunden! Nach 11.400 Jahren ist nur noch ein Viertel vorhanden usw.
Die eigentliche Altersbestimmung eines Fossils gelingt nun leicht, indem man den verbliebenen 14C-Gehalt misst. Aus dem Mengenverhältnis von 12C zu 14C kann leicht das Alter bestimmt werden!
Einschränkungen der 14C-Methode:
- Das Fossil muss noch genügend Kohelnstoffreste enthalten (Knochen,Leder, Haut, Haare usw.).
- Da die HWZ nur ca. 6000 Jahre beträgt, ist die Genauigkeit der Messung bei älteren Fossilien beeinträchtigt. Man kann heute maximal 60000 Jahre alte Fossilien zurückdatieren.
Möchte man weiter zurückdatieren, muss man andere (langlebigere) Isotope verwenden. Es eignen sich Uran- und Kaliumisotope. - Methode ungeeignet für sehr viele sehr alte Fossilien (Saurier).
- Langfristig schwankte der CO2-Gehalt der Atmosphäre. => Der 14C-Gehalt schwankt ebenfalls! Fossile Eisproben aus dem ewigen Eis der Polkappen (und anschließende Gasmessungen) können allerdings hierüber zum Teil Auskunft geben.
- Beim Messen können heutige 14C-Isotope zu Verunreinigungen führen.
- Verfälschungen durch Atombombentests
- Verfälschungen durch Freisetzen von CO2 aus Erdöl (dieses enthält logischerweise sehr viel weniger 14C).
Lösung des unglaublich schweren Wirbeltierdominos
Wiederholungsaufgaben
- Nenne zwei Gründe, die zum Aussterben einer Art führen können.
- Erkläre Ähnlichkeiten der Wirbeltierembryos im Sinne der biogenetischen Grundregel.
- Was Sind Fossilien? Nenne Beispiele
- Wie entstehen Fossilien?
- Beschreibe die 14C Methode zur Altersbestimmung.
- Die Altersbestimmung mit der 14C Methode bezeichnet man als absolute Datierung, die mit Hilfe geologischer Schichten als relative Datierung. Erkläre.
- Warum ist die folgende Aussage problematisch? „Eine 10m dicke Gesteinsschicht umfasst einen doppelt so großen Zeitraum, wie eine 5m dicke Gesteinsschicht.“