- Details
- Zugriffe: 65256
Kapitel 12.03: Evolutionstheorien

Quelle Bild: Public Domain: Original Buchtitel von Charles Darwins Buch „The Origin Of Species“
http://commons.wikimedia.org/wiki/Image:Origin_of_Species_title_page.jpg
Evolutionstheorien
Mehrere griechische Philosophen der Antike hatten in ihren Überlegungen eine Entwicklung des Lebens bereits erkannt. Aber die wichtigsten von ihnen, Platon (427-347 v. Chr.) sowie sein Schüler Aristoteles (384-322 v. Chr.), hatten eine gegenteilige Meinung. Sie prägten damit für 2000 Jahre europäische Denkweisen. Die später entstandenen jüdisch-christlichen Religionen haben diese Gedanken mit aufgenommen und in ihren Schöpfungs-“berichten“ festgehalten.
Untermauert wurden diese kreationistischen Meinungen z.B. durch Cuvier (1769-1832) einem wichtigen französischen Paläontologen und Zoologen. Er war Urheber der Katastrophentheorie, nach welcher einige wenige Katastrophen zum Aussterben vieler Arten geführt habe, was so fossile Funde erklären sollte.
Kurios ist dabei z.B., dass man auch das Alter der Erde einfach festsetzte. Der englische Bischof James Usher entschied, dass die Erde am 28. Oktober 4004 v. Chr. um 9 Uhr morgens entstanden ist. ;-)
Erst durch Lamarck und Charles Darwin und ihre ersten Theorien begann allmählich ein Wandel der Vorstellungen. Heute gilt die „synthetische Evolutionstheorie“ als die aktuelle.
Allerdings darf man hier nicht Theorie und Entwicklung verwechseln, was in der Bevölkerung, gerade auch von Kreationisten, gerne getan wird: Während es heute keinen Zweifel mehr an der Entwicklung der Arten und der Evolution an sich gibt, ist die Erklärung dieses Vorganges nach wie vor eine Theorie, da die ablaufenden Vorgänge sehr komplex sind.
Allerdings wurden weder die Darwin‘sche noch die synthetische Theorie jemals widerlegt.
Evolutionstheorien I - Lamarcks Theorie der Entwicklung der Arten
Die ersten wissenschaftlichen Evolutionstheorien stammen von Jean-Baptiste de Lamarck (1744-1829) und Charles Darwin. Der Pariser Lamarck war Mitglied der Pariser Akademie und gleichzeitig Professor für Zoologie. Im Naturhistorischen Museum in Paris beobachtete er viele Exponate als Verantwortlicher der Wirbellosensammlung. Er verglich viele Tiere, z.B. Schnecken und Muscheln und zog so seine Schlüsse.

Er erkannte als erster den Wandel der Arten und bemerkte, dass sich die Arten der Lebewesen voneinander wegentwickeln, was zu immer neuen Arten führe. Er fand aber keine heute noch vertretene Erklärung dafür. Seine Erkenntnisse veröffentlichte er 1809 in seinem Buch „Philosophie Zoologique". Seine Leistung ist also im Grunde das Erkennen der Entwicklung der Arten und der Versuch einer kausalen Erklärung.
Quelle Bild: Public domain by Wikicommonsuser Mu; http://commons.wikimedia.org/wiki/File:Jean-Baptiste_de_Lamarck.gif
Lamarckismus 1809 (aktiver Prozess!):
Theorie der Vererbung von Merkmalen nach erworbener Anpassung:
1. Alle Lebewesen besitzen eine vis vitalis (innere Körperkraft, inneres Vervollkommnungsstreben), welche Lebewesen optimal an neue Umweltbedingungen anpasst.
2. Angepasste Organe werden durch Gebrauch weiterentwickelt, bei Nichtgebrauch verkümmern diese.
Alltagserfahrungen, die Lamarcks These scheinbar stützen:
- Sonne bräunt Haut
- Muskelbetätigung führt zu Muskelwachstum.
- der lange Hals der Giraffen
- die lange Zunge des Ameisenbären
3. Die so (individuell) erworbenen Eigenschaften (=Modifikationen) werden an die Nachkommen vererbt. So liegt eine zielgerichtete Anpassung an die Umwelt vor (Das Tier HAT sich angepasst.).
Kritik an Lamarcks Theorie:
Heute weiß man, dass Modifikationen nicht vererbbar sind. Bei seinem Beispiel mit der „Vererbung“ der Muskeln eines Schmieds an seinen Sohn, hätte ihm auch auffallen müssen, das die Schwielen an den Händen des Schmieds nicht vererbt werden. ;-)
Bei vielen Arten gibt es hingegen in einem Gebiet verschiedene Ausprägungen => z.B. Löwenzahn (Tieflandform / Hochlandform)
Charles Robert Darwin

Charles Darwin, 1868 Quelle Bild: Public Domain, Julia Margaret Cameron (England, 1815-1879) 1868 http://www.photography-museum.com/darwinlg.html http://commons.wikimedia.org/wiki/Image:Charles_Darwin_by_Julia_Margaret_Cameron.jpg
Der Brite Charles Robert Darwin (12.02.1809 - 19.04.1882) ist bekannt geworden durch seine in den ersten Jahren sehr umstrittene Theorie der Abstammung. Er gilt als einer der bedeutendsten britischen Naturwissenschaftler überhaupt.
Darwin erkannte, dass es eine sehr langsame, aber stetige evolutive Entwicklung aller Organismen gibt, welche zu einer fortführenden Aufspaltung der Arten führt. Als Ursache erkannte er die Notwendigkeit der Lebewesen, sich an ihren Lebensraum anzupassen, um zu überleben.
Seine Theorie veröffentlichte er am 24. November 1859 in seinem Hauptwerk „On the Origin of Species“ (Die Entstehung der Arten). Diese Theorie bildet die Grundlage der modernen Evolutionstheorie. Sie war der entscheidende Wendepunkt in der Geschichte der modernen Biologie.
Die Grundlage für seine Theorie legte Darwin auf einer fünf Jahre dauernden Weltumsegelung mit einem Team verschiedener Wissenschaftler. Er bekam dazu, weil ein Freund ihm einen Platz auf den Forschungsschiff „Beagle“ vermittelte. Er hatte so die Chance an einer 5-jährigen Forschungsreise teilzunehmen und so viele Länder zu bereisen.
Er besuchte Südamerika, Australien und Afrika. Seine Beobachtungen hielt er durch unzählige Zeichnungen und Tagebücher fest. Auf den Galapagos-Inseln bemerkte er u.a. die enorme Formenvielfalt der dort heimischen Arten. Viele seiner bemerkenswerten Beobachtungen konnte er erst Jahre später erklären. Zum Glück war er ein guter Zeichner, sodass er viele der beobachteten seltenen Tierarten zumindest für eine spätere Auswertung zeichnen konnte.
Er wertete in den der Reise folgenden Jahren nun seine Zeichnungen und Notizen aus. Er wusste von Lamarcks Theorien und wurde ebenfalls beeinflusst durch die Aktualitätshypothese des Geologen Charles Lyell. (Die Kräfte, welche die Erde heute verändern auch schon während der gesamten Erdgeschichte am Werk waren!). Darwin zögerte lange seine Ergebnisse zu veröffentlichen. Er wusste um die Brisanz und fürchtete die Reaktion der Kirche.
Als 1858 Alfred Russel Wallace „On the variety to depart indefinitly from the original type" (über die Tendenz der Arten sich unbegrenzt weiterzuentwickeln) veröffentlichte, zog Darwin eine eigene Veröffentlichung in Betracht.
1859 veröffentlichte er dann 1250 Exemplare von „On the origin of species by means of a natural selection or the preservation of favored races in struggle for life", welches innerhalb von Stunden ausverkauft war. Es folgte natürlich eine heftige Diskussion, da die Kirche nicht akzeptieren konnte, dass der Mensch affenähnlliche Vorfahren hat - und das, obwohl Darwin den Menschen und seine Herkunft nicht in seinem Buch erwähnt hat.
Dem Menschen und seiner Abstammung widmete sich Darwin erst 8 Jahre später, im Jahr 1867 in dem Buch „Die Abstammung des Menschen und die geschlechtliche Zuchtwahl".
Seine Abstammungslehre ist unter dem kaum noch verwendeten Begriff „Darwinismus“ bekannt. Er sollte nicht mit „Sozial-Darwinismus“, einer faschistischen Ideologie des Dritten Reiches verwechselt werden.
Zusatzinformationen:
http://de.wikipedia.org/wiki/Galapagos
http://de.wikipedia.org/wiki/Charles_Darwin
http://commons.wikimedia.org/wiki/Charles_Darwin
http://de.wikipedia.org/wiki/Sozialdarwinismus
Die Weltreise auf der Beagle (1831–1836)
Ursprünglich wollte Darwin nach dem Studium eine Reise zur Insel Madeira unternehmen. Er bekam aber die Möglichkeit sich als Begleiter für Robert Fitzroy, den Kapitän der HMS Beagle, vorzustellen und so zu einer fünfjährigen Expedition (27.12.1831 - 2.10.1836) rund um den Globus aufzubrechen. Das Ziel der Expedition waren im Süden des Südamerikanischen Kontinents Patagonien und Feuerland um dort die Küsten zur Kartierung zu vermessen. Als Vorbereitung verbrachte Darwin, der einige Vorlesungen in Geologie im Studium belegt hatte, 1831 einige Wochen auf geologischen Exkursionen mit Adam Sedgwick in Nord-Wales.
Am 27. Dezember 1831 stach dann die HMS Beagle von Devonport (Plymouth) aus in See. Die Reise führte über die Kapverden, Bahia und Rio de Janeiro im Juli 1832 nach Montevideo. Die ersten zwei Jahre führte die Beagle vor allem die gewünschten Vermessungen im Gebiet zwischen Kap Horn und den Falkland-Inseln durch. Während der ersten zwei Jahre konnte Darwin an fünf Expeditionen an Land durch.
1834 bis 1835 führte Darwin weitere Expeditionen in Chile und Peru durch. Dann erreichte er schließlich mit der Beagle die Galapagosinseln.
Nachträglich auf seine Reise angesprochen, welches die beeindruckendsten Erlebnisse waren, so nannte er neben den auf Galapagos gefundenen Arten auch die Begegnung mit den Indianern Feuerlands auch die Fossilienfunde in Patagonien. Nach seiner Rückkehr nach England, analysierte Darwin ab 1836 die über 3000 mitgebrachten Exponate, Skizzen und Fundstücke.
Darwin bemerkte:
- Ähnlichkeiten zwischen Fossilien und noch lebenden Arten.
- Insbesondere fiel ihm auf, dass jede Insel ihre eigenen Schildkröten- und Vogelarten hatte, sich zwar ähnlich waren, aber von Insel zu Insel insofern (leicht) unterschieden, dass sie in Aussehen und Ernährungsweise Unterschiede aufwiesen.
Anhand von Beobachtungen der verschiedenen Tauben-, Schildkröten- und Finkenarten der Galapagosinseln entwickelte er die Theorie, dass diese jeweils von einer Ursprungsart abstammen mussten. Es gab einige Wissenschaftler seiner Zeit, die ihn durch ihre Arbeiten beeinflussten und dazu führten, dass er seine Theorie erst so formulieren konnte, wie er es schließlich tat. Groß war der Einfluss von William Paleys und seiner „Natural Theology“. Auch Lyells „Principles of Geology“ und Thomas Robert Malthus‘ Arbeiten waren einflussgebend. Malthus z.B. stellte die These auf, das die menschliche Bevölkerung schneller wächst als die Nahrungsmittelproduktion und das Wachstum durch Hungersnöte in Grenzen gehalten wird.
Zusatzinformationen:
http://en.wikipedia.org/wiki/Galápagos_Islands
http://es.wikipedia.org/wiki/Islas_Galápagos
http://commons.wikimedia.org/wiki/Atlas_of_the_Galápagos_Islands
http://commons.wikimedia.org/wiki/Category:Animals_of_the_Galápagos_Islands
http://commons.wikimedia.org/wiki/Category:Galápagos_Islands
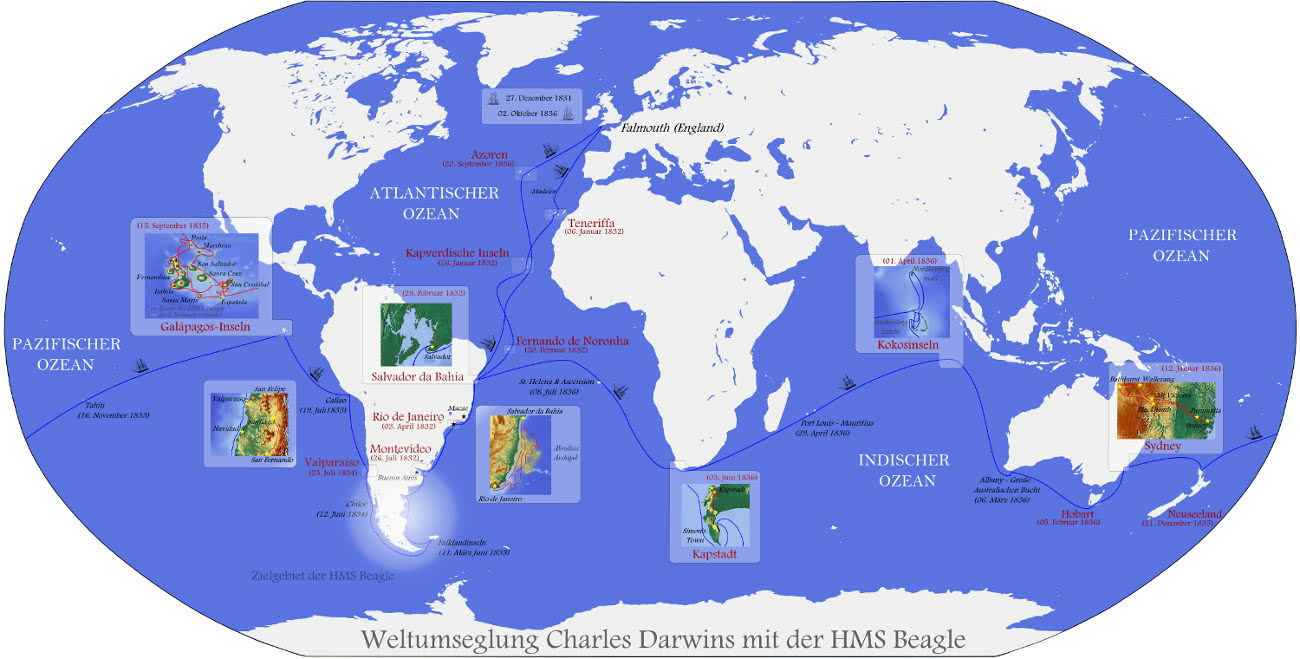
Quelle Bild: Creative Commons Attribution 3.0 Unported License by Wikicommonsuser - Devil_m25
http://de.wikipedia.org/wiki/Bild:Charles_Darwins_Weltumseglung_mit_der_HMS_Beagle_1831-1836.png - Thank You!
Die Entstehung von Darwins Evolutionstheorie
Durch seine aufmerksamen Beobachtungen der Natur bemerkte Darwin schon früh, das sich Lebewesen einer Art unterscheiden. Diese Variationen (heute spricht man von Modifikationen) sind nie völlig gleich.
Auch beobachtete er, dass fast alle Arten eine Überproduktion an Nachkommen haben und die Populationen über längere Zeiträume betrachtet dennoch konstante Zahlen haben. In Verbindung mit der Tatsache, dass Platz- und Nahrungsressourcen in jedem Ökosystem irgendwann begrenzt sind, schloss er daraus, dass die Mitglieder einer Art einen Kampf um diese knappen Ressourcen (Kampf ums Dasein - struggle for life) durchführen.
Betrachtet man nun beide Punkte zusammen, die Variabilität und den Kampf ums Dasein, so überlebt nun das Tier, welches die günstigste Variation darstellt, also am besten angepasst und somit das fitteste Tier ist (survival of the fittest). Es wird auch die meisten Nachkommen haben, da es am längsten lebt. Nach und nach kommt es so zur Veränderung der Arten, vor allem, wenn sie die äußeren Selektionsfaktoren ändern.
Von Lamarcks Theorie hatte Darwin übrigens schon früh gehört. Er erkannte zumindest die Aussage an, dass eine Vererbung von Eigenschaften stattfindet. Ob es allerdings für erworbene Eigenschaften genauso gilt, hielt er für fraglich!
Während viele Wissenschaftler der damaligen Zeit nach und nach die Logik seiner Theorie erkannten, tat sich die Kirche sehr schwer damit. Angeblich war auch Karl Marx so von Darwins Theorie begeistert, dass er sein Buch „Das Kapital“ ihm widmen wollte. Darwin lehnte dies aber ab. Auch Friedrich Engels und Thomas Henry Huxley waren Anhänger Darwins Theorie. Der deutsche Biologe und Forscher Ernst Haeckel war der erste, der Darwins Theorien (noch vor Darwin selbst) auch auf den Menschen bezog.
Die zu Ecuador gehörenden Galapagos Inseln

Quelle Bild: Public Domain von Wikipediauser CarolSpears & Earth Observatory 8270 and NASA GSFC, NOAA and MODIS - thank you
http://en.wikipedia.org/wiki/Image:Galapagos-satellite-esislandnames.jpg

Quelle Bild: CC-BY-SA-2.5, by Wikicommonsuser Michaël Lejeune, Merci http://commons.wikimedia.org/wiki/Image:Galapagos.jpg
Evolutionstheorie II - Darwins Theorie von der natürlichen Auslese (1859)
Darwin beschrieb in seinem 1859 veröffentlichtem Buch „The Origin Of Species“ (Über die Entstehung der Arten durch natürliche Zuchtwahl) den „Kampf der Arten ums Dasein“.
Dazu stellte er 5 Punkte auf, welche auf seinen weltweiten Beobachtungen basierten:
1. Überproduktion an Nachkommen: Arten produzieren in ihrer natürlichen Umwelt mehr Nachkommen, als zum eigentlichen Erhalt der Art notwendig wären.
2. Variabilität der Nachkommen: Die Nachkommen unterscheiden sich in ihrem Erbgut (Ursache sind Mutationen). Grundsätzlich sind alle Mitglieder einer Art unterschiedlich (z.B. in den phänotypischen Merkmalen). Heute weiß man, dass die Grundlage für diese Veränderungen die Mutationen der DNA sind, welche durch Rekombination verteilt werden.
3. Selektion durch die Umwelt: Einige Nachkommen sind besser an ihrer Umwelt angepasst. Ursache ist die Wirkung abiotischer und biotischer Faktoren auf die Lebewesen. Es kommt unter den Nachkommen zu einer natürlichen Auslese. Die Lebewesen, welche durch ihre jeweiligen Merkmale (zufällig!) am besten an die Umweltbedingungen angepasst sind, haben die größten Überlebens- und Fortpflanzungschancen (die größte Fitness).
=> Kampf ums Dasein (struggle for life).
=> Die am besten angepassten Individuen überleben diesen Kampf (survival of the fittest).
4. Vererbung der günstigeren Merkmale:
Es findet eine Vererbung der Merkmale statt, welche eine besonders gute Anpassung an die Umwelt erlauben. Lebewesen mit diesen Merkmalen pflanzen sich vermehrt fort und geben so die überlegenen Erbanlagen an die Nachkommen weiter.
=> Weniger angepasste und somit „untaugliche“ Lebewesen sterben aus.
5. Aktualitätsprinzip (nach Lyell):
Dieser Vorgang fand nicht nur in der Vergangenheit statt, sondern auch heute: „Die heute wirksamen Faktoren müssen auch in der Vergangenheit gewirkt haben."
=> Darwins Theorie beschreibt einen passiven Prozess!
Diese „natürliche Auslese“ führt dazu, dass Lebewesen sich den natürlichen Umweltbedingungen immer besser anpassen.
Im Verlauf langer Zeiträume führt diese Anpassung schrittweise zum Wandel der Arten.
=> Die Entwicklung geht vom Einfachen zum Komplexen
Zusatzinformationen:
http://de.wikipedia.org/wiki/Darwinismus
Evolutionstheorie III - Die Synthetische Evolutionstheorie (aktuell)
Durch neuere biologische Wissenschaften (wie Genetik, Verhaltensbiologie und Ökologie) ist heute Darwins Theorie zur „synthetischen Evolutionstheorie“ erweitert wurden. Der Begriff der „synthetischen Evolutionstheorie“ geht dabei auf J.S.Huxley den Enkel des berühmten Th.H.Huxley zurück, der ihn 1887 erstmals verwendete.
Zusatzinformationen:
http://de.wikipedia.org/wiki/Synthetische_Evolutionstheorie
1. Evolutionsfaktoren als Ursachen des Artenwandels
Heutige Evolutionstheorien beruhen auf Darwins Überlegungen. Diese müssen allerdings mit den Ergebnissen der klassischen Genetik (Mendel) und der Molekulargenetik ergänzt werden.
Basierend auf Darwins Überlegungen hat man Evolutionsfaktoren definiert:
Evolutionsfaktoren: die genetische Variabilität (Mutationen & Rekombination von Genen), Gendrift, Selektion, Isolation (sowie Domestikation).
2. Genetische Variabilität (Variation)
Für die Vielfalt der Lebewesen innerhalb einer Art sind zwei Faktoren ausschlaggebend:
2.1 Mutation des Erbguts:
- Spontane, ungerichtete Veränderung des genetischen Materials.
- Mutationen erweitern den Genpool.
- Die Mutationsrate ist bei gleichbleibenden Bedingungen recht konstant:
- Bakterien 10-7 Mutationen /Gen
- Mensch 10-6 Mutationen /Gen (das entspricht jeder 10. Keimzelle)
Präadaptation: zufällige Anpassungen an zukünftige Umweltbedingungen.
2.2 Rekombination:
Es entstehen durch geschlechtliche Fortpflanzung (bei der Meiose) ständig Neukombination von bereits vorhandenem Erbgut. So können z.B. auch rezessive Merkmale hervortreten (=> Bereitstellung neuer Genotypen).
Gene kommen in zufälliger Weise durch geschlechtliche Fortpflanzung (bei der Meiose) in neuen Kombinationen zusammen. Unter Umständen können sie so dann auch im Phänotyp erkennbar werden.
Einfluss der Rekombination:
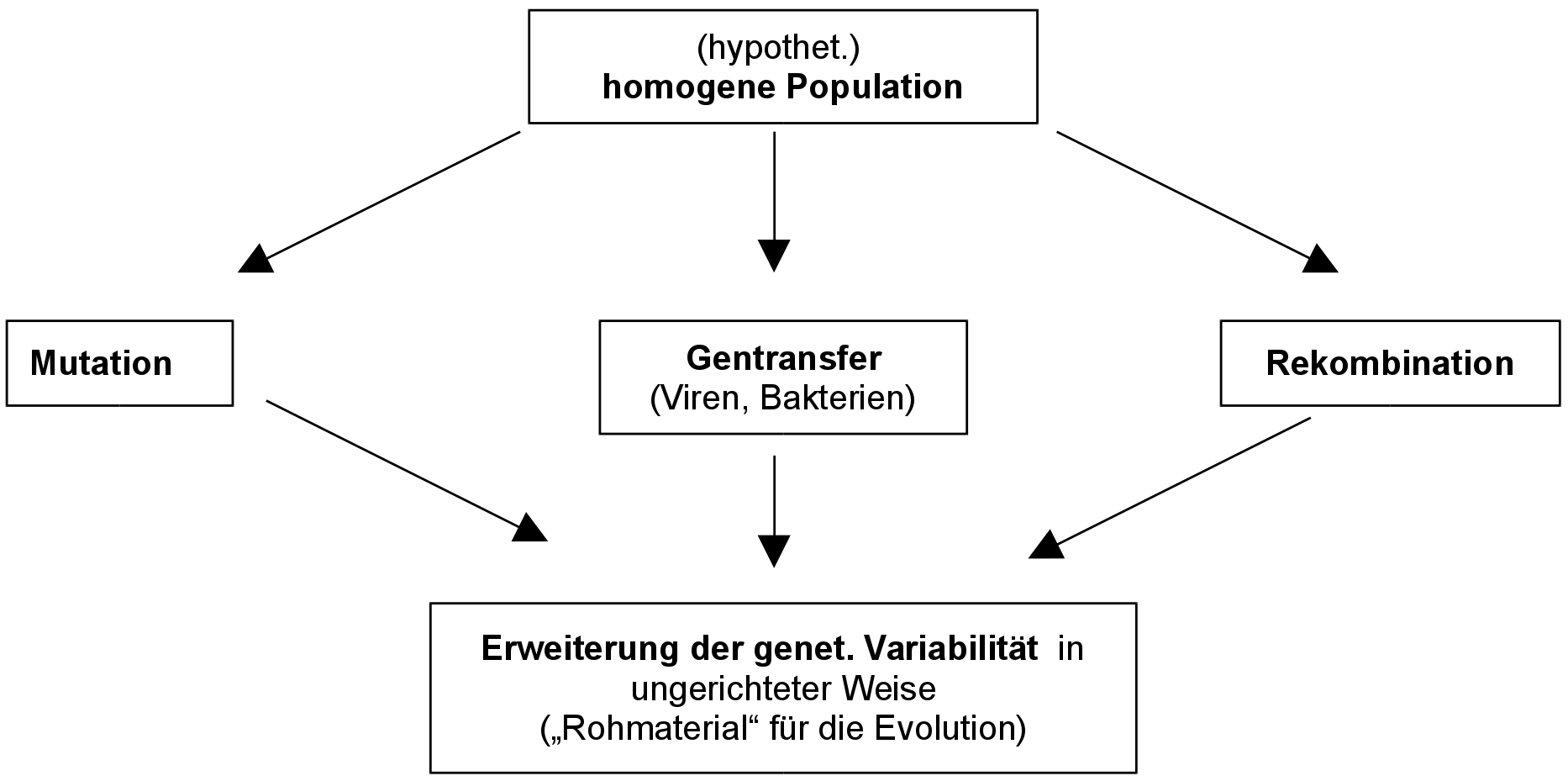
=> zufällige, ungerichtete Ereignisse stellen genetisches „Rohmaterial“ für die Evolution zur Verfügung.
3. Gendrift (Sewall-Wright-Effekt)
Um zu verstehen, was Gendrift ist, muss man zunächst den Begriff der Genhäufigkeit (bzw. der Genfrequenz) verstehen.
Genfrequenz = Häufigkeit bestimmter Gene (Allele) in einer Population.
Treten zufällig neue Gene innerhalb eines Genpools auf, so ist dies ja zunächst nur bei einem Tier.
- hohe Genfrequenz bei altbewährten Genen
- niedrige Genfrequenz bei dem neuen Gen
Als Gendrift bezeichnet man die Veränderung von Genfrequenzen (=Genhäufigkeiten) durch zufällige Auswahl von Genotypen.
Beispiel: Biologen fanden eine auffallende Grünfärbung beim Seitenfleckenleguan, der auf Inseln im Golf von Kalifornien lebt. Seine Verwandten auf dem Festland im westlichen Teil der USA haben hingegen die üblichere gelbbraune Musterung. Ganz offensichtlich wurde die Inselpopulation von wenigen Tieren gegründet.
=> Die Verschiebung von Genhäufigkeiten spielt vor allem in kleinen Populationen eine Rolle, da sich dort Genänderungen viel schneller durchsetzen können.
Dies trifft zu bei:
- Neubesiedlungen eines Gebietes durch einige wenige Gründerindividuen (zufällige Auswahl von Genotypen => Genverlust oder Genanreicherung)
- zufälliger Auswahl von (Überlebenden) Genotypen nach Naturkatastrophen (Orkane, Vulkanausbrüche, Überschwemmungen usw.).
Allgemein gilt: Je kleiner die Elternpopulation und je weniger Nachkommen produziert werden, desto stärker ist der Gendrift und somit die ständige Veränderung der Genfrequenzen.
In zu kleinen Populationen (im Extremfall bei Inzucht) setzen sich besonders negative Merkmale (wie Erbkrankheiten) durch.
4. Selektion (= natürliche Auslese)
Während Mutation, Rekombination und Gendrift Zufallsereignisse sind, kann die Selektion (natürliche Auslese) als richtender Evolutionsfaktor angesehen werden.
Selektion ist ein richtender Evolutionsfaktor!
Durch die natürliche Auslese werden die Lebewesen begünstigt, welche die günstigsten Erbanlagen für den jeweils selektierenden Faktor haben. Weniger gut angepasste Lebewesen sterben aus oder wandern ab.
Die Selektion setzt somit immer „am Phänotypen an“. Sie bewirkt dadurch eine gerichtete Verschiebung der Genhäufigkeiten. Rezessive Merkmale, welche am Phänotyp nicht erkennbar sind, werden demzufolge auch nicht selektiert.
4.1 Fitness und Darwin-Fitness
Die relative Fitness ist hoch (geringe Sterblichkeit, hohe Fertilität, gute geschlechtliche Zuchtwahl, lange Generationenlänge).
Der Begriff der biologischen Fitness kann aus dem englischen Wort „fitness“ (=Angepasstheit, Tauglichkeit) abgeleitet werden (nicht verwechseln mit der sportlichen Fitness!).
Er beschreibt den Anpassungs- bzw. Adaptationswert eines Lebewesens und somit die individuelle Überlebensrate.
Darwin-Fitness: Darwin hat eine hohe Fitness durch eine hohe Anzahl an fortpflanzungsfähigen Nachkommen angesehen. Er folgerte, dass Lebewesen mit Fortpflanzungserfolg besonders gut an ihre Umwelt angepasst sind.
Als relative Fitness bezeichnete Darwin die Fitness eines Individuums nicht direkt anhand der Gesamtanzahl seiner Nachkommen. Er setze stattdessen die Zahl der direkten Nachkommen mit dem durchschnittlichen Fortpflanzungserfolg anderer Individuen in Beziehung. Es handelt sich also im Grunde um eine mathematische Analyse der Fitness
Als „Fitness im erweiterten Sinne“ bezeichnet man die Fähigkeit eines bestimmten Genotyps, möglichst häufig im Genpool der nächsten Generation vertreten zu sein.
Das heißt, dass gut angepasste Lebewesen eine hohe Fitness haben und somit ihren Genotyp in größerem Maße an die Folgegeneration vererben als Individuen, die schlecht angepasst sind.
=> Genotypen von Lebewesen mit hoher Fitness nehmen in ihrer Häufigkeit im Genpool der Population stärker zu.
=> Die Verbesserung der Fitness ist eine der Triebkräfte der evolutiven Veränderungen der Arten.
Die direkte Fitness drückt den Anpassungs- bzw. Adaptationswert einer Art an ihre Umwelt aus. Sie beschreibt neben der relativen Überlebensrate eines Individuums auch die Häufigkeit seines Genotyps im Genpool der jeweiligen Population.
Eine hohe direkte Fitness liegt bei Tieren vor, welche viele Nachkommen haben.
Die indirekte Fitness berücksichtigt auch die Nachfahren der eigenen Nachfahren sowie die der näheren Verwandten.
4.1 Man unterschiedet drei Selektionstypen:
Stabilisierende Selektion:
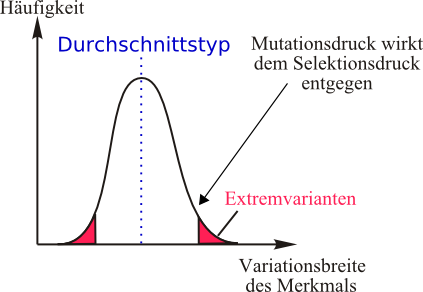
Lebewesen, welche sich bezüglich ihrer abiotischen und biotischen Faktoren im Optimum befinden, habe die meisten Nachkommen. Sie haben also eine hohe Fitness. Folglich geben sie ihre „Durchschnittsgene“ bevorzugt weiter und so sind auch ihre Nachkommen wieder ein Durchschnittstyp. Dies nennt man stabilisierende Selektion. Es wird sich immer der Durchschnittstyp durchsetzen, da er die meisten Nachkommen hat. Lebewesen, welche durch Mutation neue Gene/ Eigenschaften haben, können diese kaum weitergeben, da sie weniger Nachkommen haben.
Man findet diese Art der Selektion vor allem bei konstanten Umweltbedingungen, wenn die Lebewesen einer Art über viele Generationen hinweg schon unter diesen konstanten Umweltbedingungen leben.
(Vergleiche auch mit der „Toleranzkurve eines Organismus“ aus dem Kapitel Ökologie - abiotische Faktoren).
Individuen, welche am ehesten dem Durchschnittstyp (=Mittelwert) in Bezug auf selektierende Umweltfaktoren entsprechen, zeigen eine höhere Fitness. Vom Phänotypmittelwert stark abweichende Phänotypen können sich nicht durchsetzen.
=> Durch stabilisierende Selektion wird der Durchschnittstyp gefördert.
=> Extremvarianten werden „ausgemerzt“
=> geringe phänotypische Vielfalt.
Selektionsdruck: Stärke, mit der unterlegenere Genotypen (je nach Umweltanpassung) ausgemerzt werden.
Mutationsdruck: Tendenz zum Schwinden der Gleichförmigkeit des Erbguts.
Der Mutationsdruck ist von der Mutationsrate abhängig. Mutationen sorgen für immer neue Genotypen.
In einer hinreichend großen Wildpopulation wirkt der Selektionsdruck dem Mutationsdruck entgegen.
Richtende (auch transformierende, dynamische oder direktionale) Selektion:
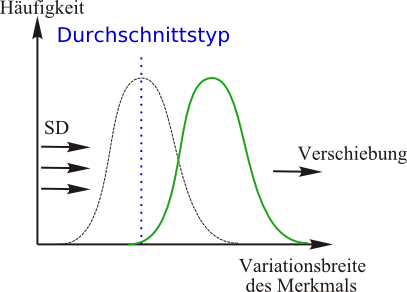
- Wenn sich Umweltbedingungen verändern, führt dies langfristig dazu, dass die am besten angepassten Lebewesen den neuen Durchschnittstypen bilden. Mittelfristig bewirken also veränderte Umweltbedingungen eine gezielte Verschiebung in Richtung der nun günstigeren Eigenschaft.
- Durch Variabilität innerhalb einer Art liegt eine große Spannbreite an Merkmalen vor.
- Lebewesen mit Phänotypen, welche ursprünglich am Rande des Merkmalsspektrums waren, werden nun durch eine Umweltänderung begünstigt (allerdings muss das Lebewesen mit dem neuen Merkmal bereits vor der Umweltänderung (hier SD = Selektionsdruck) angepasst gewesen sein (=Präadaption).
- Dieses Lebewesen hat einen Selektionsvorteil.
- Die Folge ist eine Veränderung des Genpools.
- Langfristig findet so eine ständige Anpassung der Arten an sich ändernde Umweltbedingungen statt (Artenwandel ohne Artaufspaltung).
- Menschliche Züchtung in der Landwirtschaft und bei Haustieren ist eine sehr starke richtende Selektion.
Disruptive Selektion:
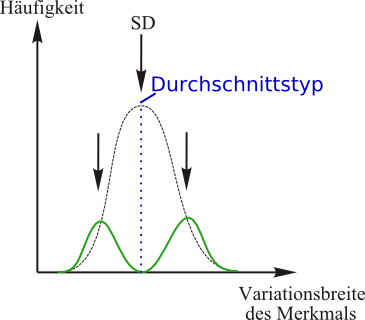
Aufspaltung der Population in zwei Rassen. Ursache ist, dass der Durchschnittstyp plötzlich in seiner Häufigkeit stark abnimmt (z.B. durch eine Krankheit oder Parasitenbefall innerhalb einer Population).
Die Extremvarianten am Ende des phänotypischen Spektrums besitzen unter Umständen besonders seltene phänotypische Merkmale (=Polymorphismus), welche es ihnen erlauben nun durch diesen Selektionsdruck eine neue ökologische Nische zu besetzen. Sie sind selektionsbegünstigt, da sie sich nun z.B. eine andere Nahrungsquelle erschließen können oder durch besonderes Verhalten dem Selektionsdruck entgehen können.
Das Besetzen neuer ökologischer Nischen durch Bildung neuer Arten (durch disruptive Selektion) wird adaptive Radiation genannt.
=> Adaptive Radiation: Auffächerung einer Stammart in neue Arten durch das Besetzen neuer ökologischer Nischen (divergente Einnischung)
Das bekannteste Beispiel für adaptive Radiation ist die Bildung der Darwinfinken auf Galapagos (siehe dazu auch Isolationsmechanismen).
Zusatzinformationen:
http://de.wikipedia.org/wiki/Selektion_%28Evolution%29
http://de.wikipedia.org/wiki/Selektionsdruck
4.2 Selektionsfaktoren I - Abiotische Selektionsfaktoren
- Temperatur:
Im Sommer 1947 nach dem extrem kalten und langen Winter 1946/47 hat man in Deutschland beobachtet, dass sich der prozentuale Anteil an großen Maulwürfen stark verringert hatte. Kleinere Maulwürfe gab es aber nach wie vor.
Was waren die Ursachen? Die geringe Temperatur hat viele im Boden lebende Insekten und Würmer erfrieren lassen, sodass nur wenig Nahrung vorlag. Diese Nahrungsverminderung wurde besonders von kleineren Maulwürfen überlebt, da diese insgesamt weniger Nahrung brauchen.
=> In diesem Fall brachte die geringe Körpergröße einen Selektionsvorteil.
=> Vergleiche dazu auch wieder die Bergmann‘sche Regel bzw. Allen‘sche Regel.
Bergmann’sche Regel: Vertreter einer Tiergruppe sind in kalten Regionen
größer als in warmen
Allen’sche Regel: Die Körperanhänge vergleichbarer Tierarten sind in
kälteren Regionen kleiner
- Wind:
Auf den Kerguelen, einer Inselgruppe in der Arktis, hat man flugunfähige Insekten- und Schmetterlingsarten gefunden. Ursache dafür, dass sich diese seltene (auch am Festland in geringem Maße vorkommende) Mutante durchgesetzt hat, könnte der starke Wind und die Kälte sein, da diese an großen Flügeln eine bessere „Angriffsfläche“ haben. Vermutlich würden beflügelte Insekten auf das Meer geweht werden.
- Licht
- Wasserversorgung
- Salzgehalt des Wasser
- Bodenverhältnisse
- Gifte (z.B. Antibiotika)
- uvm.
vgl. auch Konvergenzerscheinungen
=> Fischgestalt (Leben im Wasser)
=> Sukkulenz bei Pflanzen
4.3 Selektionsfaktoren II - Biotische Selektionsfaktoren
4.3.1 Drei interspezifische biotische Selektionsfaktoren
1. Innerartliche Selektion durch Fressfeinde (Räuber-Beute-Beziehung)
Bsp.: - Industriemelanismus beim Birkenspanner
helle Form = Normalvariante
dunkle Form = Mutation
1846 GB: einige dunkle
1895 GB: ca. 95% dunkle
- ebenso Entstehung von Mimese und Mimikry
Mimese: Nachahmung von unbeachteten Objekten in Farbe, Form und Haltung (TARNUNG)
(z.B. Stabheuschrecke, wandelndes Blatt)
Mimikry: harmlose, ungefährliche Tiere ahmen gefährliche Tiere nach (WARNUNG)
(z.B. Schwebfliege ahmt Wespe nach)
Selektion durch artverschiedene Konkurrenten: Zwei Arten beanspruchen dieselben Umweltfaktoren => Konkurrenzauschlussprinzip.
Bsp: höhere Säugetiere (Placentatiere) verdrängen Beutetiere.
Selektion durch Krankheitserreger oder Parasiten
z.B. Sichelzellenanämie (Vorteil: Widerstandsfähigkeit gegenüber Malaria)
Zusatzinformationen:
http://de.wikipedia.org/wiki/Mimikry
http://de.wikipedia.org/wiki/Mimese
Aufgaben zu biotische Selektionsfaktoren:
1. Bestimme, welche Form vorliegt: Mimikry oder Mimese?

Stabheuschrecke


Feuersalamander

2. Erkläre begründet, warum beide Strategien funktionieren und das Überleben gegenüber Fressfeinden erleichtern.
3. Die Schmetterlinge der Gattung Dismorphia liegen besonders langsam und wären eine leichte Beute für Vögel. Obwohl sie eigentlich zu den Weißlingen gehören, ähneln sie Schmetterlingen der Gattung Ithomiini. Welche Vermutung liegt nahe?
4.3.2 Intraspezifische biotische Selektionsfaktoren
Selektion durch innerartliche Konkurrenz: gleiche Umweltbedingungen.
Turnier- und Kommentkämpfe: keine ernsthafte Verletzung der Artgenossen, aber dennoch Ermittlung des Stärkeren (z.B. Hirsche).
Selektion durch geschlechtliche Zuchtwahl (sexuelle Selektion).
Sexualdimorphismus: Männchen und Weibchen sind phänotypisch verschieden (z.B. Prachtgefieder, Geweih, etc.).
=> Verstärkung von Generation zu Generation
Aber: Unter Umständen können solche Merkmale aber auch einen Selektionsnachteil darstellen (bunte Tiere sind für Räuber besser sichtbar oder auch bei der Flucht zu langsam, da das Geweih schwer ist usw.! - Der Säbelzahntiger sowie der Riesenhirsch starben vermutlich wegen einer solchen Übersteigerung aus).

Dammwild

Rothirsch (Rotwild)
4.4 Künstliche Auslese als besondere Form der Selektion
Menschen betreiben vermutlich seit ca. 10000 Jahren die Zucht von Tieren. Man nennt die so gezüchteten Tiere auch domestizierte Lebewesen. Sie werden als Haustiere oder Nutzpflanzen verwendet.
- Der Mensch wählt als Züchter bestimmte Individuen aus der Wildpopulation aus (=> Nur ein kleiner Teil des Genpools wird ausgewählt). Die Selektion erfolgt nach den gewünschten Phänotypen (z.B. hohe Milchleistung bei Kühen, süßes Aussehen bei Hunden, große Hühnereier, schönes Fell bei Katzen, usw...)
- Der gewünschte Populationsteil wird isoliert und veränderten Selektionsbedingungen ausgesetzt.
- Durch Domestikation werden seltene Mutationen, die sich vermutlich in freier Wildbahn nicht halten könnten, ausselektiert und bevorzugt und kontrolliert gepaart.
- Diese künstliche Selektion (Zuchtwahl) wirkt im Grunde der natürlichen Auslese entgegen.
- Die Folgen sind:
- morphologische Veränderungen (z.B. Hängeohren)
- Leistungssteigerungen (z.B. Milchkuh)
- Verhaltensänderungen (z.B. bei Verkürzung des Gesichtsschädels bei Hunden, Instinktreduktion)
- Stoffwechseländerungen (z.B. Reduktion des Fettes bei Schweinen,)
Die auffälligste Folge der Domestikation ist die auftretende (und zunehmende) Formenvielfalt gegenüber den ursprünglichen Wildformen. Der Grund liegt vermutlich darin, dass in einer hinreichend großen Wildpopulation der Selektionsdruck dem Mutationsdruck entgegenwirkt.
Selektionsdruck: Stärke, mit der bestimmte, unvorteilhafte Genotypen (je nach Umweltanpassung) ausgemerzt werden.
Mutationsdruck: Tendenz des Erbgutes zur permanenten Veränderung und somit zum Schwinden der Gleichförmigkeit des Erbguts.
Zusatzinformationen:
http://de.wikipedia.org/wiki/Synthetische_Evolutionstheorie
http://de.wikipedia.org/wiki/Evolution_%28Systemtheorie%29
http://de.wikipedia.org/wiki/Selektion_%28Evolution%29
5. Entstehung der Artenvielfalt durch Isolation
Isolation ist ein Faktor, welcher nicht ausschließlich eine räumliche Trennung einer Art beschreibt (diese nennt man Separation), sondern auch andere Barrieren einschließt, welche zu einer Teilung des Genpools führen. Die Teilpopulationen unterliegen dann unterschiedlichen Selektionsdrücken und entwickeln sich unabhängig voneinander. Am Ende der Isolation ist eine Fortpflanzung von je einem Individuum der voneinander getrennten Gruppen nicht mehr möglich. Zwei Arten haben sich gebildet (=allopatrische Artbildung).
Barrieren können neben unterschiedlichen Verhaltensweisen bei der Balz auch geographische Barrieren wie Ozeane, Gebirge, Flüsse, Autobahnen, Eiszeiten und Wüsten sein. Entstehen sie im Laufe der Zeit neu (z.B. durch Kontinentaldrift), kann eine Population gespalten werden. Es kann zu einer unterschiedlichen Entwicklung der beiden Populationen kommen.
5.1 Artkonstanz und Artenwandel
- Stabilisierende Selektion => Artkonstanz
Das Aussehen der Art verändert sich nicht oder kaum. - Dynamische Selektion => Artenwandel
Verschiebung des Aussehens bzw. anderer Merkmale.
= allochrone Artbildung => keine Vermehrung der Arten.
- Disruptive Selektion => Aufspaltung der Population in zwei Rassen
= synchrone Artbildung => Artaufspaltung
5.2 Aufspaltung der Arten
Der Gendrift, Seewall-Wright-Effekt, führt theoretisch zu einer gleichmäßigen Durchmischung des Genpools.
Panmixie: Andauernde, gleichmäßige Durchmischung des Erbguts innerhalb einer Population. D.h. für alle Individuen besteht die Möglichkeit der Paarung in gleicher Weise.
=> homogene Population; gleichmäßige Verteilung der Allele
Aber in der Realität: höhere Wahrscheinlichkeit für Paarung zwischen Verwandten (wg. räumlicher Nähe)
- genetische Unterschiede zwischen Teilpopulationen
- Auftrennung des Genpools
Initialvorgang für die Artaufspaltung:
Separation = räumliche Trennung einer Population in Teilpopulationen
=> Verhinderung der Panmixie.
Zusatzinformationen:
http://de.wikipedia.org/wiki/Allopatrische_Artbildung
http://de.wikipedia.org/wiki/Sympatrie
http://de.wikipedia.org/wiki/Sympatrische_Artbildung
5.3 Isolationstypen
5.3.1 Geographische Isolation (=Separation)
In Europa kommen zwei sehr ähnliche Spechtarten vor, der Grünspecht und der Grauspecht. Beide haben einen gemeinsamen Vorfahren und sind eng miteinander verwandt. Obwohl sie in verschiedenen ökologischen Nischen leben (der Grünspecht lebt eher am Boden und sucht dort Insekten, wie Ameisen, der Grauspecht hingegen pickt am Baum die Larven und Insektennahrung aus dem morschen Holz) und sie fast komplett getrennte Genpools haben, sind dennoch Mischzüchtungen gelungen.
=> Beide stellen verschiedene Arten dar, die Genpools sind aber noch nicht sehr lange voneinander getrennt.
Aufgrund der Anzahl an Mutation pro Generation kann man ungefähr zurückrechnen, wann beide Arten einen gemeinsamen Vorfahren hatten. Dies war in der letzten Eiszeit vor ca. 100 000 - 65 000 Jahren der Fall.
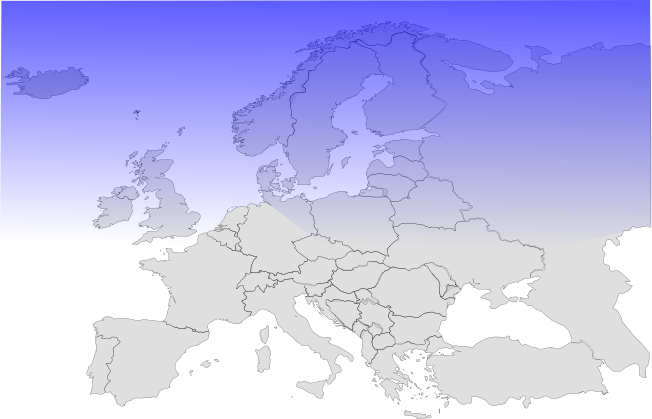
Die Eiszeit vor 65000 Jahren. Der blaue Bereich war mit Schnee dauerbedeckt.
Die Eiszeit separierte die Population der ursprünglichen Vorfahren in zwei Gruppen. Eine lebte vorwiegend im westlichen Teil Europas (iberische Halbinsel), die andere im südöstlichen (Türkei). Denn auch die Bereiche, welche nicht durch das ewige Eis bedeckt waren, boten wenig Nahrung, da ca. bis auf Höhe der Alpen keine Wälder mehr zu finden waren (Steppe). Die beiden Teilpopulationen waren nun getrennt und über viele Jahrtausende fand kein Kontakt zwischen den beiden (nun kleineren) Genpools statt. In kleineren Genpools können sich Mutationen schneller ausbreiten und zwar nur im eigenen Genpool, nicht in dem der isolierten Verwandten.
Da beide Populationen nun auch verschiedenen Selektionsbedingungen unterlagen, kamen nach und nach andere Phänotypen zum Vorschein. Je länger die Trennung dauerte, desto verschiedener wurden beide Populationen. Es kam zur Rassenaufspaltung, später zur Artspaltung.
Ist die Artspaltung manifestiert, z.B. durch verschiedene Paarungszeiten, Paarungsriten oder durch das Nichterkennen des Partners durch unterschiedliche Balzfärbungen, so kann auch keine erneute Zusammenführung der Arten stattfinden.
So kam es am Ende der Eiszeit dazu, dass beide Arten nun wieder zum großen Teil in einem gemeinsamen Gebiet vorkommen, aber die Arten sich nicht wieder vermischt haben.
5.3.1.1 Was führt zur geographischen Isolation:
1. Großflächige Gebiete
Eine Population bewohnt ein großes Gebiet. Die Grenzen des Gebietes liegen weit auseinander. Die große Entfernungen zwischen den Randgebieten kann zu vielfältigen Mutationen, Selektionsfaktoren und Gendrift führen.
Beispiele: britische Heringsmöwe – amerikanische Silbermöwe – britische Silbermöwe – skandinavische Silbermöwe (entstanden aus der asiatischen Stammart).
2. Gründerpopulationen
Einzelne (u.U. trächtige) Lebewesen werden in neue, aber isolierte Gebiete getragen (z.B. durch einen Sturm, Meeresströmungen oder Überschwemmungen). Dort können sie eine Neubesiedlung durchführen (exponentielles Wachstum), allerdings ist der ursprüngliche Genpool sehr klein, sodass Mutationen eine sehr hohe Wahrscheinlichkeit haben, sich durchzusetzen.
3. Klimatische Veränderungen
Ganze Populationen werden durch Eiszeit oder Wüstenbildung in andere Gebiete gedrängt und dabei separiert (siehe Beispiel Grün- und Grauspecht).
4. Landveränderungen größeren Ausmaßes
Trennung durch Lavaströme, Kontinentaldrift, Senkung des Landes oder Ansteigen des Meeresspiegels usw.
Geographische Separation ist die wichtigste Voraussetzung für eine divergente Entwicklung neu aufgespaltenen Teilgenpools.
1. Jede Teilpopulation hat nur einen Teil des Genpools! (=> unterschiedliche Genfrequenzen => Gendrift)
2. In den Teilpopulationen kommen unterschiedliche (zufällige) Mutationen vor.
3. In den beiden Lebensräumen herrschen verschiedene abiotische und biotische Faktoren, sodass es zu unterschiedlicher Selektion kommt.
=> Die Separation wirkt in unterschiedlicher Richtung (= disruptive Selektion).
Der erste Schritt einer solchen Auseinanderentwicklung ist nach vielen Jahrtausenden die Entstehung von zwei verschiedenen Rassen (= geographische Subspezies).
Eine Aufhebung der Separation durch Kontakt der Teilpopulationen, hat bei verschiedene Rassen die Folge, dass es im Gebiet des Aufeinandertreffens zur Verbastardierung („Bastardierungszone“) kommt (siehe z.B. Nebelkrähe - Saatkrähe).
- verschiedene Arten keine Folgen. Es findet keine Verbastardierung mehr statt. Die Arten sind voneinander getrennt.
Zusatzinformationen:
- http://de.wikipedia.org/wiki/Eiszeitalter
- http://de.wikipedia.org/wiki/Grauspecht
- http://de.wikipedia.org/wiki/Grünspecht
- http://de.wikipedia.org/wiki/Bild:Picus_viridis_dis.png (Lebensraum Grünspecht)
- http://de.wikipedia.org/wiki/Bild:Picus_canus_distr_new.png (Lebensraum Grauspecht)
5.3.1.2 Beispiele für erfolgte Aufspaltungen durch geographische Isolation
a) Evolution der Möwenarten
In der Vergangenheit unseres Planeten kam es immer wieder zu Eiszeiten, bei denen Tiere aus ihrer ursprünglichen Heimat in weiter im Süden liegende Gebiete verdrängt wurden.
So auch im Quartär, als ein Großteil der skandinavischen Silbermöwen in ein Gebiet rund um das kasparische Meer ausweichen mussten. Am Ende der Eiszeit konnten die Möwen ihre alten Territorien wieder zurückerobern. Allerdings gab es nun auch andere Möwenarten. An den Randgebieten der jeweiligen Territorien kommt es auch immer noch zu Bastardierungen. Nur zwischen der britischen Heringsmöwe und die britische Silbermöwe können keine gemeinsamen Nachkommen bilden.
1. Erklären Sie, warum die beiden britischen Möwenarten keine Rassenbastarde bilden können.
2. Stellen Sie eine nachvollziehbare Hypothese zur Verbreitung und zur Vielfalt der heutigen Möwenarten auf.
3. Erklären Sie, inwieweit hier das Konkurrenzvermeidungsprinzip Anwendung findet.
b) Evolution der Krähenarten
In Westeuropa gibt es ausschließlich schwarze Krähen. In Osteuropa hingegen gibt es ausschließlich grau-schwarze Krähen, die sogenannten Nebelkrähen.



Aufspaltung der Krähenarten während der Eiszeit

Nebelkrähe

Beispiele für geographische Isolation durch den Menschen:

Autobahnen und Flüsse durchtrennen Ökosysteme und teilen Populationen

5.3.2 Ökologische Isolation
Das bekannteste Beispiel für eine ökologische Isolation sind die Darwinfinken der Galapagosinseln (der eine geographische Isolation vermutlich vorausging). Man findet auf diesen Inseln 14 endemische (heimische, auf ein Verbreitungsgebiet beschränkte) Arten.
Merkmal einer ökologischen Isolation ist die spezielle Einnischung der Arten als Anpassung an unterschiedliche Jahreszeiten, Tageszeiten oder andere abiotische Faktoren.
Man geht also nicht von einer Auftrennung durch eine räumliche Trennung der Arten, sondern von einer ökologischen Auftrennung aus. Merkmal ist das Erschließen neuer ökologischer Nischen.
Entstehungstheorie der Darwinfinken:
Verdriftung der körnerfressenden Vorfahren, welche bodenlebend waren, vom Festland auf die Insel:
=> starke Vermehrung, da auf Galapagos gute Bedingungen vorlagen und v.a. keine Fressfeinde vorhanden waren
=> zunehmende Nahrungskonkurrenz, da es vorerst zu einem exponentiellen Wachstum, dann zur Nahrungsknappheit am Boden kam (da ja alle Bodenfinken dieselben Umweltfaktoren beanspruchen).
=> Schon vorhandene Varianten (Präadaptation) mit anderen Schnabelformen können auf andere Nahrung ausweichen (welche nach wie vor im Überfluss vorhanden ist - z.B. Kakteen) und haben somit plötzlich einen Selektionsvorteil.
=> Diese Varianten besetzen nun die neuen ökologischen Nischen und entziehen sich dadurch dem innerartlichen Konkurrenzkampf der anderen Artgenossen.
=> Nach und nach spalten sich die Nachfahren der Varianten vom Genpool ab. Neue Rassen entstehen (später dann neue Arten).
Adaptive Radiation: Auffächerung einer Stammart in neue Arten durch das Besetzen neuer ökologischer Nischen (divergente Einnischung)
Man kann die adaptive Radiation im Grunde in zwei Typen unterteilen:
Allopatrische Artbildung:
Die Artaufspaltung beruht auf geographischer Isolation (= Separation).
Sympatrische Artbildung:
Die Artaufspaltung findet im gleichen Verbreitungsgebiet statt (z.B. durch disruptive Selektion).
Ein weiteres Beispiel für die adaptive Radiation ist die Verbreitung der Beuteltiere in Australien.
Übersetzung:
- allos = anders
- patria = Heimat
- sym = mit, zusammen
5.3.3 Reproduktive Isolation
Können zwei Lebewesen keine Nachkommen mehr zeugen, obwohl das Genom dies noch erlauben würde, so spricht man von einer reproduktiven Isolation. Die Ursachen können dafür vielfältig sein:
Begegnung von Männchen und Weibchen
↓
unterschiedliche Lebensräume beider Tiere
↓
Beginn der Balz durch spezielle Auslöser (AAM)
↓
Falsche Signale lösen keine Balz aus.
↓
Handlungskette der Balz
↓
keine korrekte
„Antwortreaktionen“
auf Signale des Partners
↓
Paarung
↓
unpassende Geschlechtsorgane
↓
Befruchtung der Zygote (Eizelle)
↓
unpassendes Erbgut (z.B. stimmt die Chromosomenzahl?)
↓
Entwicklung zur Blastula, Morula, Fetus
↓
Steuern die Hormone die Entwicklung richtig?
↓
Geburt
Passt der Nachwuchs durch den Geburtskanal?
↓
Nachkommen
Sind die Nachkommen steril?
Sind sie sonst benachteiligt?
↓
Schwein gehabt ;-)
Noch liegt eine Art vor!
=> keine reproduktive Isolation
Man kann alle Gründe, welche eine reproduktive Isolation bewirken, in folgende Gruppen zusammenfassen:
- Räumliche Isolation:
Die Tiere treffen einfach nicht aufeinander,
z.B. können Tiger und Löwe fruchtbare Nachkommen zeugen, sie begegnen sich aber nicht.
- Ethologische Isolation:
Veränderung im Ablauf des Balzverhaltens (=> unterschiedliche Handlungsketten) oder auch der Auslöser der Balz (AAM) hat sich verändert.
- Mechanische Isolation:
Die Kopulationsorgane der Tiere passen nicht erfolgversprechend zusammen.
- Jahreszeitliche Isolation:
Unterschiedliche Paarungszeiten führen zu wenig Erfolg (manchmal auch zu Migräne ;-)).
z.B.: paart sich und brütet die britische Silbermöwe drei Wochen früher als die britische Heringsmöwe.
- Unverträglichkeit des Genoms:
Zygotenabsterben durch nicht zueinander passendes Genom,
z.B: Schaf + Ziege können keinen Nachwuchs haben, da der Embryo abstirbt.
- Verminderte Vitalität der Bastarde:
Sterilität der Bastarde (Bsp. Hase/ Kaninchen ; Maulesel/Maulpferd)
Die Folgen dieser Störungen sind:
a) Individuen können sich nicht mehr paaren.
b) Es entstehen Bastarde, welche deutliche Selektionsnachteile haben!
- schlechtere Anpassung an die ökologische Nische als die reinrassigen Nachkommen
=> Ausselektion durch Konkurrenzdruck - weniger deutliche ausgeprägte Sexualmerkmale wie die reinrassigen Nachkommen
=>Probleme bei der Partnerfindung => Keine Weitergabe der Gene. - Sterilität (Maulesel/Maultier (=Maulpferd))
Bei Pflanzen gibt es weitere Besonderheiten:
- Nicht jeder Pollen bildet auf jeder Narbe einen Pollenschlauch aus (chemische Inhibition).
- Bestäubende Insekten fliegen meist nur auf die Blüten einer Variante, von der sie „wissen“, dass sie sicher Nektar trägt.
- Einige Pflanzen haben so seltsam geformte Blüten, dass nur angepasste Insekten bestäuben können. Diese bleiben ihrer Art ebenfalls „treu“.
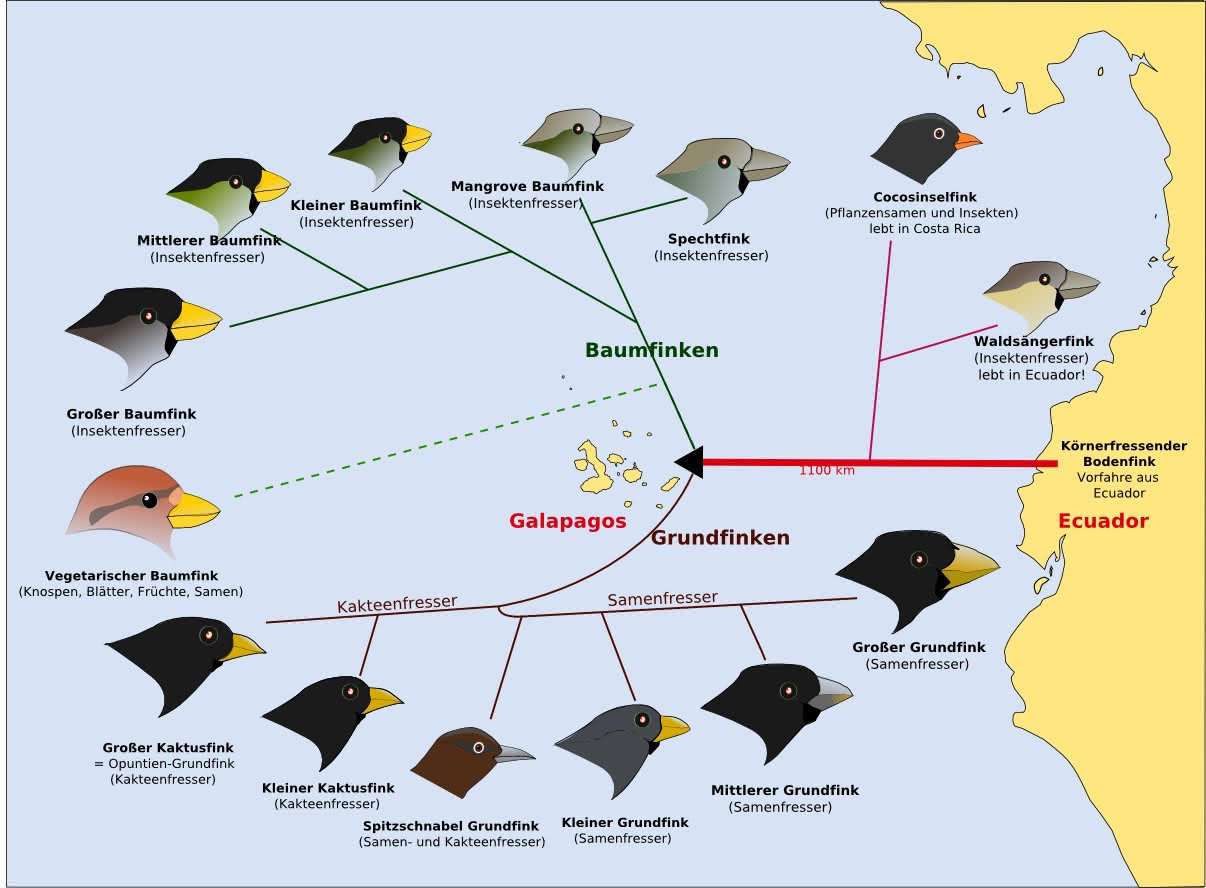
Die Adaptive Radiation am Beispiel der Darwinfinken
Adaptiv = anpassend
Radiation = Strahlung, Ausbreitung
Darwin kam 1835 nach Galapagos. Der Begriff „Darwinfinken“ beschreibt die Gruppe der auf Galapagos von Darwin beschriebenen Singvogelarten (und dem Kokosfinken (Pinaroloxias inornata), welcher nur auf der vor Costa Rica liegenden Cocos-Insel vorkommt).
Die 14 sehr eng verwandten Singvogelarten stammen alle von einem gemeinsamen Vorfahren ab, welcher übrigens kein Fink (so wie man allgemein glaubt!) war.
Neuere genetische Untersuchungen haben gezeigt, dass der gemeinsame Vorfahr ein finkenähnlicher Singvogel, aus der Gattung der Tangaren (Thraupidae) (verwandt mit den Ammern (Emberizidae)) war.
Tipp: Den Begriff adaptiv kennst Du bereits aus dem Kapitel Sinnesorgane - Auge. Die Adaptation ist die Anpassung der Linse in ihrer Brechkraft.
Die 14 Darwinfinkenarten unterteilen sich in die folgenden vier Gruppen:
1. Grundfinken (Geospiza)
- Groß-Grundfink (auch Dickschnabel Grundfink) (Geospiza magnirostris)
- Mittel-Grundfink (Geospiza fortis)
- Klein-Grundfink (Geospiza fuliginosa)
- Spitzschnabel-Grundfink (G. difficilis)
- Kaktus-Grundfink (G. scandens)
- Opuntien-Grundfink (G. conirostris)
2. Baumfinken (Camarhynchus)
- Dickschnabel-Darwinfink (C. crassirostris)
- Mangrove-Darwinfink (C. heliobates)
- Papageischnabel-Darwinfink (C. psittacula)
- Zweig-Darwinfink (C. parvulus)
- Kleinschnabel-Darwinfink (C. pauper)
- Spechtfink (C. pallidus)
3. Certhidea-Finken
- Waldsängerfink (C. olivacea)
4. Pinaroloxias-Finken
- Kokosfink (P. inornata)
Evolution im Kleinen:
Darwin kam 1835 nach Galapagos. Das Landesinnere bietet wenig Futterquellen. Manchmal fällt monatelang kein Regen. Um 1977 herum fiel auf der Insel Daphne (in der Mitte des Archipels) zum Beispiel gar kein Regen. Durch die Dürre starben 85% der Finken. Es überlebten vor allem die Finken mit großen Schnäbeln, welche die harten Samen fressen konnten, die noch vorhanden waren.
Als 1983 durch das Phänomen El Niño es 8 Monate durchgehend regnete und sich das Meer um 4°C erwärmte, starben viele Fische und Meerestiere. Viele Vögel wanderten ab. Auf der Insel Daphne stieg die Temperatur und es wurde durch das Wasser sehr grün. Einige wenige Grundfinken kamen nach Daphne und brüteten dort. Nach nur 15 Jahren haben sie sich auf mehrere 100 Paare vermehrt. Von Generation zu Generation wurden die Schnäbel großer. Die Finkenschnäbel waren besser an die dortige Nahrung angepasst.
Adaptive Radiation der „Darwinfinken“ in der Übersicht:
Zusatzinformationen:
http://de.wikipedia.org/wiki/Geospiza_fortis ;
http://de.wikipedia.org/wiki/Spitzschnabel-Grundfink
Finkenarten auf Galapagos:
Jede Finkenart ist auf eine Nahrungsart spezialisiert. Demzufolge sieht man bei den Finken besonders große Unterschiede an den Schnäbeln. Die Ernährung ist Ursache für die unterschiedlichen Körperanpassungen. Dies sieht man besonders in den verschiedenen Schnabelformen.
14 Arten:
- Spechtfink sucht Käferlarven am Holz der Bäume an Kratern. Manchmal benutzt er einen kleinen Zweig als Werkzeug, um besser an die Larven zu kommen.
- Der kleine Grundfink sammelt harte Samen vom Boden
- der Mittelgrundfink frisst Beeren
- der große Grundfink frisst harte große Samen
- der große Kaktusfink frisst Kaktusfleisch der Opunzien
- Dickschnabelfink frisst weiche Samen
- Laubsängerfink fängt Spinnen und Insekten
- manche Finken fressen Hautparasiten von großen Echsen (Drusenköpfe)
Der Mittelgrundfink (Geospiza fortis):
Dieser Vogel verfügt über einen festen Schnabel, mit dem sich kleinere Samen gut öffnen lassen. Findet er keine kleinen Samen, z.B. in Jahren, in denen durch Trockenheit nur wenige Früchte gebildet wurden, versucht er, größere Samen zu knacken, Beeren zu fressen oder Insekten zu fangen.
Selbst die Hautparasiten der Leguane können eine Nahrungsquelle für ihn sein (Symbiose). Die Leguane lassen dies zu, weil sie so die lästigen Parasiten loswerden.
Im Grunde konnte er also alle Nahrungsquellen, welche heute von anderen Darwinfinkenarten besetzt sind, nutzen. Vermutlich war er also eine frühe Entwicklungsphase im Prozess der sympatrischen Artbildung. Es gibt von ihm zwei „Morphen“, welche im gleichen Gebiet leben. Sie unterschieden sich in ihrer Schnabelgröße und paaren sich bevorzugt mit Tieren der gleichen Schnabelgröße.
Ursache für diese Präferenz bei der Partnerfindung sind Unterschiede im Gesang durch die unterschiedlichen Schnabelgrößen.
Der Spitzschnabel-Grundfink (Geospiza difficilis):
Dieser Vogel hat seine Bekanntheit als Vampirfink erhalten. Neben der Vampirfledermaus ist er die einzige Wirbeltierart, welche sich vom Blut anderer Wirbeltiere ernährt (vor allem der Vogelarten Blaufußtölpel und Nazcatölpel). Auf diese Weise deckt der Vogel auf den kleineren, wasserlosen Inseln Energie- und Wasserbedarf. Interessant ist, dass die Tölpel sich nicht dagegen wehren. Vermutlich waren die Vorfahren der Spitzschnabel-Grundfinken also Mittelgrundfinken, denen sie auch ähnlich sind, sodass die Tölpel evtl. davon ausgehen, von Parasiten befreit zu werden. Spitzschnabel-Grundfinken sind vom Aussterben bedroht!
Auf Wulf-Island gibt es Finken, die die Eier, welche von Tölpeln nicht mehr beachtet werden, da sie aus dem Nest gerollt sind weiterrollen und dann über die Klippen rollen, wo sie am Boden zerbersten.
Andere Finken picken kleine Wunden mit ihren spitzen Schnäbeln in die Ellenbogenhaut der Vögel. Vermutlich ist diese Ernährungsweise aus dem Fressen von Hautparasiten entstanden.
Zusatzinformationen:
http://de.wikipedia.org/wiki/Darwinfinken
Der Stammbaum der Darwinfinken

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Fitness und Evolution sozialer Verhaltensweisen
Honigbienen gaben den Wissenschaftlen lange Jahre Rätsel auf. Die Arbeiterinnen sind Weibchen, welche steril sind. Sie können keine Nachkommen haben. Ihre Fitness ist also = 0! Warum arbeiten die unfruchtbaren Weibchen ohne eine Chance auf Weitergabe ihrer Gene?
Ursache:
Durch diese „Selbstaufgabe“ werden nahe Verwandte geschützt, gefördert oder unterstützt.
=> höherer Fortpflanzungserfolg der Verwandten, welche ja einen Großteil der eigenen Gene haben (große Übereinstimmung!)
=> Ein Großteil der Gene des Helfers werden (selbst im Falle des Todes) in die nächste Generation eingebracht.
=> Verwandtenselektion
Ein solches Verhalten wird auch als Altruismus bezeichnet:
Altruistisches Verhalten: Gruppendienliches Verhalten, bei dem ein Tier seine eigene Fortpflanzung zugunsten von Artgenossen einstellt.
Obwohl hier ein Beispiel für eine fehlende Fitness vorliegt, kann man sehen, dass das Prinzip der Bienen sehr gut funktioniert. Biologen haben für solche Fälle den Begriff der Gesamtfitness definiert.
Gesamtfitness = Fitness durch Weitergabe der eigenen Gene durch eigenen Reproduktionserfolg + Fitness durch Weitergabe der eigenen Gene durch Verwandte
=> Biologisch betrachtet steht also nicht der Organismus bzw. das Individuum im
Zentrum der Betrachtung, sondern das Gen/ Genom.
=> „Egoismus der Gene“
Aufgabe zu den drei behandelten Evolutionstheorien


Der lange Hals der Giraffen hat nur 7 Wirbel, genau wie bei allen anderen Säugetieren. Trotzdem ist er mehrere Meter lang und ermöglicht das Fressen von Blättern in großer Höhe.
Stelle gegenüber, wie Lamarck und Darwin jeweils das Entstehen des Langen Halses bei Giraffen erklärt hätten. Gib abschließend eine begründete Erklärung, wie man es heute nach der synthetischen Evolutionstheorie erklären würde.


Der lange Hals der Giraffen kann nach Darwin oder Lamarck unterschiedlich interpretiert werden!
Wohin geht die Reise? Die Richtung der Evolution
Rekombination und Mutation sind rein zufällige Prozesse. Da aber die Entwicklung von einfachen Arten zu komplexen Arten führt, muss es richtungweisende Elemente in diesem zufälligen Geschehen geben.
Die Selektion bestimmt also die Richtung der Entwicklung.
Das wichtigste Ziel der Evolution ist damit nicht die Perfektion von Eigenschaften (Bsp. Für imperfekte Strukturen sind die Zähne bei Elefanten, der Rücken des Menschen usw.), sondern um ein Erreichen einer hohen Fitness und somit um einen hohen Fortpflanzungserfolg.
Dieser kann auf mehreren Weisen erreicht werden:
- neue, bessere Enzyme
- Erschließen neuer Nahrungsquellen (Pelzmotten leben von organischem Gewebe, was es erst durch den Menschen gab!)
- Effektivere Brutfürsorge
- Bessere Abwehr (und Resistenz) von Krankheiten und Parasiten
- Bessere Anpassung an abiotische Faktoren (Kälte, Hitze, Trockenheit, Lichtmangel usw.)
- Besserer Schutz vor Fressfeinden durch Tarnung, schnellere Flucht, Kampf usw.
Jede Verbesserung der Fitness durch neue Eigenschaften bringt potentiell den Vorteil mit sich, das mehr Nachkommen entstehen. Diese neuen Eigenschaften werden dabei dann besonders häufig vererbt.
Einfluss auf den Genpool:
Der Genpool ist der Genbestand einer Art. Er umfasst alle Allele dieser Art. Jedes einzelne Individuum dieser Art besitzt aber immer nur einen Teil des tatsächlich vorhandenen Genpools. Durch Mutationen kommen immer neue Allele hinzu!
Beide Faktoren zusammen führen zur genetische Variabilität der Arten.